
Cladogram of Cannales based on DNA sequence data.
Zingiberales Griseb., Grundr. Syst. Bot.: 167. 1-2 Jun 1854 [‘Zingiberides’]; Zingiberidae Cronquist in Brittonia 30: 505. 19 Dec 1978; Zingiberanae Takht. ex Reveal in Phytologia 74: 179. 25 Mar 1993
Fossils Spirematospermum is represented by leaves, trilocular fruits and seeds from the Campanian to the Maastrichtian and the Cenozoic of Europe and North America. The seeds are spirally striate, operculate and have a chalazal chamber and a micropylar collar. Several zingiberalean fossils have been described from the Maastrichtian Deccan Intertrappean Beds in India, such as Heliconiaites, Cyclanthodendron, Musocaulon and Heliconites, which are fossilized leaves and stems, and Musa cardiosperma, which comprises fruits with seeds, although it is questioned whether these should be attributed to Musa. Tricostatocarpon and Striatornata are trilocular fruits and operculate seeds from the Campanian of Mexico. Leaves from Late Cretaceous layers in the United States have been described as Zingiberopsis magnifolia, and some fossil seeds from the Eocene of Oregon resemble extant Ensete.
Habit Bisexual, perennial, often giant, herbs (stem rarely lignified), often with a pseudostem consisting of superimposed leaf sheaths. Sometimes with stilt roots. Rhizome and roots usually rich in starch.
Vegetative anatomy Roots usually with V-shaped aggregations of xylem, with large metaxylem element in angle. Phellogen absent. Stem cortex and central tissue with each vascular bundle surrounded by dense fibre envelope; cortex and central tissue separated by cylinder of fibres. Secondary lateral growth absent. Vessels present in roots (rarely also in stem). Vessel elements with scalariform or simple perforation plates; lateral pits? Imperforate tracheary xylem elements tracheids. Wood rays absent. Axial parenchyma? Sieve tube plastids usually P2cs type (with starch grains and cuneate protein crystals; sometimes P2cfs type, with starch grains, and cuneate protein crystals and filaments). Nodes multilacunar with several leaf traces. Stem sometimes with lysigenous mucilaginous cavities and ducts. Tanniniferous idioblasts often frequent. Idioblasts with suberised cell walls and containing aromatic (ethereal) oils and resins. Silica often present in vascular bundle sheaths. Silica bodies druse-like, stellate, conical or boat-shaped (sometimes present in stegmata), or absent. Calciumoxalate as acicular, rhomboidal, prismatic or tubular crystals, or as raphides or crystal sand. Starch grains usually simple (sometimes compound), often flat (sometimes irregular), usually excentric (sometimes compound; rarely spherical or elliptic).
Trichomes Hairs unicellular or multicellular, uniseriate or multiseriate, simple or branched (rarely furcate to stellate or candelabra-like), or absent.
Leaves Alternate (usually distichous, sometimes tetrastichous or spiromonostichous), simple, entire, often differentiated into pseudopetiole and pseudolamina, with supervolute ptyxis (laterally rolled-up in bud). Stipules absent; leaf sheath usually open (sometimes closed), with or without ligule at distal end. Pseudopetiole bundle transection arcuate. Venation pinnate-parallelodromous (with midvein, S-shaped lateral veins and fine transverse venation). Stomata usually tetracytic or paracytic, with symmetrical guard cells (rarely polycytic, hexacytic or tricytic); neighbouring cells with non-oblique divisions, or annular cells with non-oblique or oblique divisions. Cuticular wax crystalloids as platelets or longitudinally aggregated rodlets (Strelitzia type), or absent. Mesophyll with idioblasts containing ethereal oils and/or calciumoxalate as druses, prismatic or rhomboidal crystals or crystal sand (raphides usually absent). Epidermis cells often with spherical silica bodies or silica sand. Leaf margin entire.
Inflorescence Usually terminal (rarely axillary), capitate or cone-like, or simple or branched thyrse (sometimes raceme-like or spicate), consisting of few- to many-flowered (sometimes single-flowered) cymules/cincinni subtended by spiral bracts (bracts rarely absent). Partial inflorescences sometimes consisting of enantiomerous cincinni. Bracts persistent, often large. Floral prophylls (bracteoles) sometimes lateral, sometimes absent.
Flowers Zygomorphic or asymmetrical. Epigyny. Tepals 3+3; outer tepals with imbricate aestivation, usually sepaloid, free or more or less connate into tubular or spatha-like structure; inner tepals with imbricate aestivation, petaloid, more or less connate into tube; when zygomorphic, then median outer tepal abaxial. Gynoecial/septal nectaries epigynous, inserted at or sunken into ovary apex (sometimes labyrinthine); one or three glandular nectaries often present at stylar and staminal bases (sometimes also androecial nectaries on staminodia). Disc usually absent.
Androecium Stamen one, five, 3+2 or 2+3 (rarely 3+3) fertile, and one to four staminodial (rarely absent), often petaloid. Filaments free or connate at base, usually adnate to tepal base (sometimes free or absent). Anthers dorsifixed or basifixed, versatile or non-versatile, disporangiate or tetrasporangiate, introrse, usually longicidal (dehiscing by longitudinal slits; rarely poricidal, dehiscing by pores). Tapetum secretory or amoeboid-periplasmodial. Staminodia one to five, variously arranged, often petaloid, free or more or less connate, sometimes fused with fertile stamen (sometimes absent).
Pollen grains Microsporogenesis usually successive (sometimes simultaneous). Pollen grains usually inaperturate (sometimes monosulcate, disulcate, sulcatoporate, 5–16-pantoporate, or spiraperturate), shed as monads, bicellular or tricellular (rarely unicellular) at dispersal. Exine tectate, with columellate infratectum, or intectate, consisting of scattered columellae or spinules, verrucate, gemmate, echinate, scabrate, rugulate, striate or smooth, or exine absent.
Gynoecium Pistil composed of usually three (rarely two) eusyncarpous or paracarpous connate carpels; median carpel abaxial. Ovary inferior, usually trilocular (later often incompletely or entirely trilocular or unilocular, rarely bilocular). Style single, simple, filiform, hollow, inserted in groove in filament of fertile stamen and between thecae. Stigma infundibuliform, cupular, decurrent or trilobate (rarely bilamellate), papillate or non-papillate, Wet type. Pistillodium absent.
Ovules Placentation axile, basal or subbasal (when ovary trilocular) or parietal (when ovary unilocular), rarely basal due to reduction of septa. Ovules four to more than 50 per carpel, anatropous or orthotropous (finally sometimes amphitropous), apotropous, bitegmic, crassinucellar or weakly crassinucellar. Micropyle usually endostomal (sometimes bistomal or exostomal). Cylindrical thickening, ’micropylar collar’, well developed? Parietal tissue one or two cell layers thick, formed from archesporial cell (sometimes absent). Nucellar cap often present. Epidermal cells of megasporangium often radially elongate and forming ‘nucellar pad’. Megagametophyte monosporous, Polygonum type. Antipodal cells ephemeral, not proliferating. Endosperm development ab initio nuclear or helobial; chalazal chamber often early degenerating. Endosperm haustoria? Embryogenesis asterad, caryophyllad or chenopodiad.
Fruit Usually a loculicidal, dry or fleshy, capsule or a drupe (sometimes an irregularly dehiscing capsule, or a nut; rarely a berry or a syncarp with drupaceous or baccate mericarps).
Seeds Aril usually present. Seed coat endotestal. Exotesta consisting of fibriform elongate cells and often with phytomelan layer on epidermal cell walls (sometimes lignified or sclerenchymatous). Mesotesta sclerotic, tanniniferous or absent. Endotestal cells with silica, usually sclerotic, often with thickening U-shaped in cross-section, or absent. Operculum formed from inner testal epidermis, present near radicula and combined with ‘micropylar collar’ developed from outer integument and forming annular invagination in perisperm surrounding operculum. Tegmen thin, undifferentiated, often crushed. Perisperm well developed, rich in compound starch grains. Chalazosperm well developed or absent. Endosperm usually sparse, with aleurone (sometimes with lipids and/or starch grains). Embryo usually straight (sometimes curved or plug-shaped), often well differentiated, without chlorophyll or lipid bodies, Trillium type. Cotyledon one, ligulate. Cotyledon hyperphyll compact, not assimilating. Hypocotyl internode short to long. Mesocotyl absent. Coleoptile present or absent. Collar usually indistinct. Collar rhizoids sometimes present. Germination phanerocotylar? or cryptocotylar.
Cytology n = 7–14, 17, 21, 25 (x = 11)
DNA Plastid gene atpA with 6 bp deletion. Triplication of nuclear CYC-like gene. Duplications of GLOBOSA-like genes.
Phytochemistry Flavonols (kaempferol, quercetin, myricetin, isorhamnetin, syringetin), O-methylated flavonols, flavonol sulfates, dihydroflavones, flavone-C-glycosides, flavanones, luteolin, chalcones, cyanidin, delphinidin, pelargonidin, malvidin, etc., anthoxanthins, deoxyanthocyanins (apigenin etc.), ethereal oils consisting of mono-, di-, tri- and sesquiterpenes, proanthocyanidins (prodelphinidin, procyanidin), phenylpropanoids and related curcumins (diarylheptanoids), caffeic acid, chelidonic acid, zingerone (phenol), indole alkaloids, quinines, glycosides (e.g. 3-glycoside, 3-rutenoside), cyanogenic glycosides, steroidal saponins, resins, aromatic acids and ketones, benzoic quinones, shikimic acid- or polyacetate-derived arthroquinones, phenylphenalenones, and oxypipe colanic acid (polysaccharide). Cell walls with ferulic acid? Ellagic acid not found.
Systematics Zingiberales are sister-group to Commelinales with strong support.
The basal branching in Zingiberales is still unclear, although Musaceae are sometimes identified (with low support) as sister to the rest. Heliconia (Heliconiaceae) is part of the basal polytomy. Strelitziaceae and Lowiaceae appear to be sister-groups. According to Stevens (2001 onwards) these share the following potential synapomorphies: pseudopetiole with adaxial and abaxial series of air canals; perianth whorls coloured, distinct; adaxial stamens of inner whorl sterile; floral column (sterile apex of ovary) present; stigma trilobate; aril hairy; and exostomal aril lobed or fimbriate.
A well supported clade is [[Cannaceae+Marantaceae]+[Zingiberaceae+Costaceae]], which is characterized by the following features (Stevens 2001 onwards): silica bodies druse-shaped, boat-shaped; absence of raphides; pseudopetiole short and poorly differentiated, with a single series of air canals; stomatal guard cells with inner and outer ledges unequal (asymmetrical in transverse section); inner tepals connate; adaxial stamen of inner whorl fertile; tapetum amoeboid-periplasmodial, non-syncytial; both androecial whorls with two staminodia; micropyle endostomal; ‘micropylar collar’ well developed; exotestal cells longitudinally elongate; endosperm sparce; and presence of chalazosperm.
The [Cannaceae+Marantaceae] clade has the synapomorphies (Stevens 2001 onwards): pseudopetiole with oblique cells; flowers asymmetrical, pairwise; only one staminal half fertile; staminodia free; stigma not very expanded; endosperm absent or almost so; cells of chalazal intrusion into megasporangium degenerating and forming chalazal canal; and x=9.
The clade [Zingiberaceae+Costaceae] is characterized by: ligulate leaves; outer tepals connate; nectaries two, at apex of ovary; labellum formed by lateral staminodia of outer staminal whorl and by two staminodia of inner whorl; labellum large, with narrow tube and distinct open limb; exine present; style slender, running between two anthers halves; stigma cupular or infundibular; endosperm development helobial; chalazal mass in seed developed; endosperm persistent, not very abundant; and hypocotyl well developed.
|
|
Cladogram of Cannales based on DNA sequence data. |
CANNACEAE Juss. |
( Back to Cannales ) |
Genera/species 1/8–10
Distribution Tropical America.
Fossils Unknown.
Habit Bisexual, perennial herbs, often giant herbs. Rhizome rich in starch, often tuberous.
Vegetative anatomy Phellogen absent. Secondary lateral growth absent. Vessels present in roots. Vessel elements with scalariform or simple perforation plates; lateral pits? Imperforate tracheary xylem elements tracheids. Wood rays absent. Axial parenchyma? Sieve tube plastids P2cs type, with cuneate protein crystals and starch grains, without protein filaments. Nodes? Stem in some species with numerous lysigenous mucilaginous cavities and canals. Tanniniferous idioblasts abundant. Silica bodies druse-shaped or stellate, adjacent to vascular bundles. Calciumoxalate as acicular, rod-shaped or plate-like or prismatic single crystals (raphides absent). Starch grains flat.
Trichomes Hairs absent.
Leaves Alternate (usually distichous, sometimes spiral), simple, entire, with supervolute ptyxis (laterally rolled-up in bud), often differentiated into pseudopetiole and pseudolamina. Stipules absent; leaf sheath open, without ligule. Venation pinnate-parallelodromous, with distinct mid-vein and numerous longitudinal primary veins. Stomata tetracytic, hexacytic or paracytic; guard cells with inner and outer ledges uniform. Cuticular wax crystalloids as longitudinally aggregated and often extremely long rodlets (Strelitzia type). Epidermis without silica bodies. Secretory cavities absent. Mesophyll without mucilaginous idioblasts; mesophyll cells with calciumoxalate as druses or single crystals (raphides absent). Leaf margin entire.
Inflorescence Terminal, simple or compound thyrse (sometimes raceme- or spike-like), usually with two-flowered cymules, cincinni; each cincinnus a pseudanthium with two asymmetrical flowers in each pair being mirrors of each other, enantiomery. Floral prophyll (bracteole) lateral, on right side of one flower and on left side of second flower in each pair.
Flowers Asymmetrical, large. Epigyny. Tepals 3+3; outer tepals sepaloid, persistent, free; inner tepals petaloid (one smaller), connate at base into a tubular structure. Septal/gynoecial nectaries sunken into ovary apex. Disc absent.
Androecium Stamen one fertile (adaxial outer stamen), petaloid, with one theca (two microsporangia), and (one to) three (or four) petaloid staminodia one inner of which, inserted opposite stamen, forming labellum and remaining ones forming ‘wings’; median adaxial inner stamen absent. Filaments somewhat connate at base, adnate to tepals. Anther dorsifixed, non-versatile, disporangiate (one theca degenerating), introrse, longicidal (dehiscing by longitudinal slits); placentoid present. Tapetum amoeboid-periplasmodial? or secretory?, with binucleate to sexanucleate cells. Staminodia usually three (rarely one, two or four).
Pollen grains Microsporogenesis simultaneous or successive. Pollen grains inaperturate, shed as monads, tricellular? or bicellular? at dispersal. Exine intectate, consisting of scattered columellae or spinules.
Gynoecium Pistil composed of three connate carpels; median carpel abaxial. Ovary inferior, trilocular, tuberculate to muricate. Style single, simple, flattened, petaloid, together with staminodia forming floral ‘upper lip’. Stigma terminal, decurrent along one stylar margin, papillate, Wet type. Pistillodium absent. Secondary pollen presentation (pollen grains deposited on abaxial side of style prior to anthesis).
Ovules Placentation axile. Ovules c. 20 to more than 50 per carpel, anatropous, bitegmic, crassinucellar. Micropyle endostomal. Outer integument approx. ten cell layers thick. Inner integument ? cell layers thick. A cylindrical ridge-shaped thickening, ‘micropylar collar’ (‘Mikropylarkragen’), formed from outer integument surrounding micropyle. Hypostase present. Parietal cell? Megagametophyte monosporous, Polygonum type. Endosperm development ab initio nuclear. Endosperm haustoria? Embryogenesis asterad.
Fruit Usually a loculicidal capsule, tuberculate to muricate and glandular (sometimes a nut), with persistent outer tepals.
Seeds Seed pachychalazal (mitosis taking place in chalaza and basal part of megasporangium during ovule development). Aril probably as hairs from funicle; seed coat impermeable since exotesta and chalazal epidermis form a protecting layer of Malpighian-like cells; germination slit often developed adjacent to raphe, uplifted and hence permitting water uptake of seed. ‘Micropylar collar’ absent. Seed coat testal. Exotesta consisting of fibriform elongate Malpighian-like cells, without phytomelan layer. Mesotesta sclereidal. Endotesta and tegmen absent. Perisperm well developed, hard, copious, starchy. Chalazal tissue, chalazosperm, well developed. Endosperm one cell layer thick, with aleurone. Embryo straight, well differentiated, Trillium type, chlorophyll? Cotyledon one. Cotyledon hyperphyll compact, not assimilating. Hypocotyl internode short. Mesocotyl absent. Coleoptile present or absent (cotyledon sheath somewhat prolonged). Radicula well developed, persistent. Collar roots present. Germination?
Cytology n = 9 – Polyploidy occurring.
DNA
Phytochemistry Flavonols (kaempferol, quercetin), cyanidin, tannins, proanthocyanidins (prodelphinidin), chelidonic acid, and aromatic and other resins present. Ellagic acid, alkaloids, saponins, and cyanogenic compounds not found.
Use Ornamental plants, starch sources (South American or Queensland arrowroot from Canna edulis).
Systematics Canna (8–10; southeastern United States, Mexico, the West Indies, Central America, tropical and subtropical South America, with the largest diversity in northwestern South America).
The pollen grains in Canna are similar to those in its sister-group Marantaceae.
COSTACEAE (Meisn.) Nakai |
( Back to Cannales ) |
Genera/species 7/110–115
Distribution Pantropical, with their largest diversity in Central and South America, New Guinea and northern Australia.
Fossils Unknown.
Habit Bisexual, perennial (sometimes giant) herbs, often with aerial stem. Rhizome rich in starch. Root nodules rarely present. Rarely epiphytic. Rarely with axillary bulbils. Stem usually simple (in Tapeinochilos branched, with lateral branches penetrating leaf sheaths).
Vegetative anatomy Phellogen absent. Secondary lateral growth absent. Vessels present in roots (sometimes also in stems) Vessel elements with scalariform perforation plates; lateral pits? Imperforate tracheary xylem elements tracheids. Wood rays absent. Axial parenchyma? Sieve tube plastids P2cs type, with cuneate protein crystals and starch grains, without protein filaments. Nodes? Ethereal oils absent. Raphide sacs and tanniniferous cells absent. Silica as stellate, druse-like or as single bodies. Calciumoxalate crystals frequent (raphides absent).
Trichomes Hairs unicellular or multicellular, uniseriate.
Leaves Alternate (tetrastichous or spiromonostichous: spirally arranged), simple, entire, with supervolute ptyxis (laterally rolled-up in bud). Stipules absent; leaf sheath closed, with a row of adaxial air canals (air canals absent from pseudopetiole and pseudolamina) and ligule in transition zone to pseudolamina. Vascular bundles adaxial. Hypodermis at least one-layered. Venation pinnate-parallelodromous; mid-vein distinct; secondary veins parallel, connected by transverse tertiary veins. Stomata tetracytic or paracytic; guard cells with inner and outer ledges non-uniform. Cuticular wax crystalloids absent. Epidermis without silica bodies. Mesophyll without mucilaginous idioblasts; mesophyll cells with calciumoxalate as druses or single prismatic crystals (raphides absent). Leaf margin entire.
Inflorescence Usually terminal (sometimes axillary), usually dense, cone- or spike-like (flowers in Monocostus solitary, axillary). Bracts imbricate, persistent, often with abaxial extrafloral nectaries at apex. Floral prophyll (bracteole) lateral, tubular or plicate.
Flowers Zygomorphic, large. Epigyny. Tepals 3+3; outer tepals with imbricate aestivation, sepaloid, persistent, connate into a tubular perianth (sometimes spatha-like); perianth tube on one side split to base; median outer tepal abaxial; inner tepals with imbricate aestivation, petaloid, longer than outer tepals, unequal in size, connate at base into a tube; median inner tepal larger, often bent upwards. Septal/gynoecial nectaries two, sunken into ovary apex. Disc often present.
Androecium Stamen single (probably median adaxial inner stamen), often petaloid. Filament flattened, free from tepals. Anther dorsifixed?, usually inserted in middle or lower part of filament (in Tapeinochilos at apex), non-versatile, tetrasporangiate, introrse, longicidal (dehiscing by longitudinal slits); connective often apically widened (in Tapeinochilos pectinate). Placentoid present. Tapetum secretory, with binucleate cells. Staminodia five including abaxial stamen in outer whorl, petaloid, connate into a labellum inserted opposite stamen and usually much longer than perianth (labellum in Tapeinochilos strongly reduced); stamen and labellum connate at base into a tube.
Pollen grains Microsporogenesis successive. Pollen grains disulcate to sulcatoporate (foraminate; Dimerocostus, Monocostus), 5–16-pantoporate (foraminate; Costus), or spiraperturate (Tapeinochilos), shed as monads, bicellular at dispersal. Exine thick (resistant against acetolysis), consisting of rodlets packed into layers and covering interapertural areas.
Gynoecium Pistil composed of usually three (rarely two) connate carpels. Ovary inferior, usually trilocular (rarely unilocular or bilocular). Style single, simple, filiform, usually more or less adnate to filament of fertile stamen and inserted inside a groove between two anther halves (thecae); stylar canal present. Stigma usually cupular with fimbriate margins (sometimes bilamellate, consisting of two ciliate structures with a dorsally bilobate appendage), papillate, Wet type. Pistillodium absent.
Ovules Placentation usually axile (rarely parietal, then ovary unilocular). Ovules c. 15 to more than 50 per carpel, anatropous, bitegmic, crassinucellar. Micropyle endostomal. Outer integument five or six cell layers thick. Inner integument ? cell layers thick. A cylindrical ridge-shaped thickening, ‘micropylar collar’ (‘Mikropylarkragen’), formed from outer integument surrounding micropyle. Hypostase present. Epidermal cells of megasporangium vertically elongate. Parietal cell? Parietal tissue? Megagametophyte monosporous, Polygonum type (or tetrasporous, Adoxa type?). Endosperm development helobial. Endosperm haustoria? Embryogenesis caryophyllad.
Fruit Loculicidal or irregularly (rarely explosively) dehiscing capsule with persistent outer tepals (rarely nutlike).
Seeds Aril often present. Seed coat testal. Exotesta consisting of fibriform elongate cells? and with phytomelan layer on epidermal cell walls. Endotesta well developed, with cells rich in silica. Tegmen degenerated. Operculum present adjacent to radicula and combined with ‘micropylar collar’. Perisperm well developed, rich in starch (with single, concentric starch grains). Chalazal tissue, chalazosperm, well developed. Endosperm sparse, with oily lipids, without starch. Embryo large, straight, with lipid bodies, Trillium type, with chlorophyll. Cotyledon one, differentiated into one sheath-like and one lamina-like part, photosynthesizing, with apical outgrowth directed backwards. Cotyledon hyperphyll compact (small), not assimilating (present at apex of large foliaceous assimilating sheath). Hypocotyl internode long. Coleoptile absent. Collar? Germination phanerocotylar?
Cytology n = 9, 18 (Costus); n = 14 (Dimerocostus)
DNA
Phytochemistry Flavone-C-glycosides, steroidal saponins, cyanogenic compounds, and benzoquinones present.
Use Ornamental plants.
Systematics Chamaecostus (7; C. congestiflorus, C. curcumoides, C. cuspidatus, C. fragilis, C. fusiformis, C. lanceolatus, _; tropical South America), Dimerocostus (3; D. argenteus, D. cryptocalyx, D. strobilaceus; Central America, tropical South America), Monocostus (1; M. uniflorus; Amazonian Peru), Paracostus (2; P. englerianus: tropical West and Central Africa; P. paradoxus: Borneo), Hellenia (2; H. globosa: Southeast Asia, West Malesia; H. speciosa; India, Andaman and Nicobar Islands, southern China, Southeast Asia, Malesia to New Guinea, the Bismarck Archipelago and Queensland), Tapeinochilos (16–20; the Moluccas, New Guinea to tropical Australia), Costus (c 75; tropical regions on both hemispheres).
Costaceae are sister-group to Zingiberaceae.
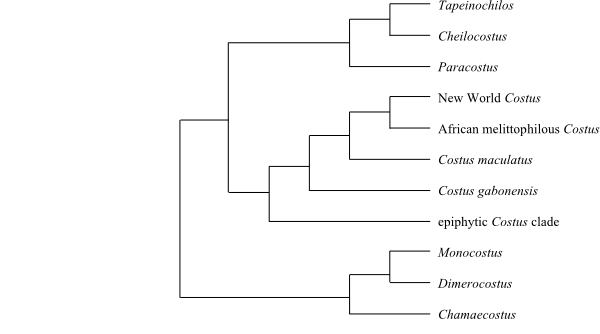 |
Cladogram of Costaceae based on morphology and DNA sequence data (Specht 2006). |
HELICONIACEAE Vines |
( Back to Cannales ) |
Genera/species 1/190–200
Distribution Tropical America, few species in East Malesia and Melanesia to Samoa.
Fossils Unknown.
Habit Bisexual, perennial (often giant) herbs. Rhizome starchy.
Vegetative anatomy Rhizome with endodermis-like layer. Phellogen absent. Secondary lateral growth absent. Vessels present in roots. Vessel elements with scalariform perforation plates; lateral pits? Imperforate tracheary xylem elements tracheids. Wood rays absent. Axial parenchyma? Sieve tube plastids P2cs type, with cuneate protein crystals and starch grains, without protein filaments. Nodes? Laticifers absent. Silica bodies boat-shaped, in idioblasts, stegmata, associated with vascular bundles. Idioblasts, raphide sacs, with calciumoxalate raphides abundant. Crystal sand present or absent. Rhizome with cylindrical or ellipsoidal starch grains.
Trichomes Hairs multicellular, uniseriate or branched (sometimes candelabra-shaped).
Leaves Alternate (distichous), simple, entire, differentiated in pseudopetiole and pseudolamina, with supervolute ptyxis (laterally rolled-up in bud). Stipules absent; leaf sheath well developed, without distinct ligule. Venation pinnate-parallelodromous, with distinct midvein and arcuate, densely spaced secondary veins, converging at apex and connected to transversal tertiary veins. Stomata usually polycytic (sometimes paracytic or tetracytic); adjacent cells with oblique divisions. Cuticular wax crystalloids as longitudinally aggregated rodlets (Strelitzia type), chemically dominated by wax esters. Epidermis without silica bodies. Mesophyll with mucilaginous idioblasts containing calciumoxalate raphides (and/or crystal sand). Leaf margin entire.
Inflorescence Terminal, spike-like thyrse consisting of few- to many-flowered cincinni, each subtended and often enclosed by large boat-shaped spathe-like green to brightly coloured and usually persistent bract. Floral bracts small and usually membranous. Floral prophyll (bracteole) keeled, lateral.
Flowers Obliquely zygomorphic (asymmetric). Epigyny. Tepals 3+3, petaloid, five tepals connate into a tubular upper lip; median outer tepal adaxial, fused with remaining tepals only at base. Septal nectaries, a triradiate apically closed crater opening by three pores at stylar base, present in axile tissue from below locules and upwards. Disc absent.
Androecium Stamens 2+3 fertile. Filaments filiform, free, adnate at base to perianth tube. Anthers basifixed, non-versatile, tetrasporangiate, introrse, longicidal (dehiscing by longitudinal slits). Tapetum amoeboid-periplasmodial, non-syncytial. Median outer stamen modified into subulate to petaloid staminodium, inserted in perianth tube opposite median outer tepal.
Pollen grains Microsporogenesis successive. Pollen grains inaperturate (functionally monoaperturate), heteropolar, shed as monads, tricellular at dispersal. Exine thin and reduced, absent or discontinuous at distal pole. Pollen surface spinulate to psilate.
Gynoecium Pistil composed of three connate carpels. Ovary inferior, trilocular. Style single, simple, filiform, hollow, often somewhat curved. Stigma one, capitate or trilobate, papillate, Wet type. Pistillodium absent.
Ovules Placentation subbasal-axile. Ovule one per carpel, anatropous, apotropous, bitegmic, crassinucellar. Micropyle bistomal. Outer integument six to nine cell layers thick. A cylindrical ridge-shaped thickening, ‘micropylar collar’ (‘Mikropylarkragen’), formed from outer integument, surrounds micropyle. Inner integument two cell layers thick. Hypostase absent. Parietal cell formed from archesporial cell, not forming a distinct parietal layer or parietal tissue early degenerating. Megagametophyte monosporous, Polygonum type. Endosperm development ab initio nuclear. Endosperm haustoria? Embryogenesis?
Fruit Usually a drupe with one pyrene per locule (sometimes a schizocarp with two or three drupaceous mericarps; sometimes on prolonged pedicel). Mechanical layer mainly formed by endocarp.
Seeds Aril absent or almost so. Micropylar operculum formed from funicle and combined with ‘micropylar collar’. Testa thin, undifferentiated. Tegmen thin, undifferentiated. Perisperm one-layered, well developed, with starch and aleurone, surrounding endosperm. Endosperm copious, starchy and oily (with single, concentric starch grains). Embryo straight or slightly curved, little differentiated, Trillium type, chlorophyll? Cotyledon one. Cotyledon hyperphyll compact, not assimilating. Hypocotyl internode short. Coleoptile absent; leaf sheath lobed. Collar at right angles to cotyledon. Germination?
Cytology n = (11) 12
DNA
Phytochemistry Insufficiently known. Tannins and proanthocyanidins present. Cyanogenic compounds not found.
Use Ornamental plants.
Systematics Heliconia (190–200; Sulawesi, the Moluccas, New Guinea, the Bismarck Archiplago, Solomon Islands, Fiji, Samoa, southern Mexico, Central America, the West Indies, tropical South America, with their highest diversity in tropical America).
The species delimitations in Heliconia are notoriously difficult.
The abaxial outer stamen is sterile, a feature similar to that in the [[Cannaceae+Marantaceae]+ [Zingiberaceae+Costaceae]] clade. Heliconia is sister-group to the remaining Zingiberales in the study by Janssen & Bremer (2004), but the support for this placement is generally low.
LOWIACEAE Ridl. |
( Back to Cannales ) |
Lowiales Takht. ex Reveal et Doweld in Novon 9: 551. 30 Dec 1999
Genera/species 1/16
Distribution Southern China, Southeast Asia, the Philippines, Borneo, islands in western Pacific.
Fossils Unknown.
Habit Bisexual, perennial herbs. Rhizome rich in starch.
Vegetative anatomy Rhizome with endoderm-like layer on inner side of cortex; outer cortex often with fibre strands. Phellogen absent. Secondary lateral growth absent. Vessels present in roots. Vessel elements with scalariform perforation plates; lateral pits? Imperforate tracheary xylem elements tracheids. Wood rays absent. Axial parenchyma? Sieve tube plastids P2cs type, with cuneate protein crystals and starch grains, without protein filaments. Nodes? Idioblasts, stegmata, with conical silica bodies in association with vascular bundles in stem and leaves. Calciumoxalate as raphides and single crystals. Rhizome cells with crystal sand. Starch grains compound.
Trichomes Hairs absent.
Leaves Alternate (distichous), simple, entire, differentiated into pseudopetiole and pseudolamina, with supervolute ptyxis (laterally rolled-up in bud). Stipules absent; leaf sheath well developed, without ligule. Pseudopetiole with adaxial and abaxial series of air canals. Venation pinnate-parallelodromous, with distinct midvein and parallel secondary veins, connected with transverse tertiary veins; longitudinal veins abruptly ascending and converging towards apex. Stomata paracytic; guard cells deeply sunken, asymmetrical (with inner and outer borders unequal). Cuticular wax crystalloids absent. Epidermis without silica bodies. Mesophyll with mucilaginous idioblasts containing calciumoxalate raphides. Leaf margin entire.
Inflorescence Terminal, cymose, consisting of repeated one- or few-flowered partial inflorescences, branching from scale-like bracts below flower. Prophyll two-keeled; above these two interphyll with axillary partial inflorescence buds; above these a bract with an axillary flower; on top a reduced bract enclosing inflorescence apex.
Flowers Zygomorphic, often resupinate and then with median inner tepal adaxial, more or less evil-smelling. Epigyny. Tepals 3+3; outer tepals sepaloid, connate at base into a tubular structure; median outer tepal adaxial; inner tepals petaloid, free; two lateral inner tepals smaller and sometimes narrow and enclosing stamens; median inner tepal modified into large sometimes boat-shaped adaxial labellum (in resupinate flowers abaxial). Septal/gynoecial nectaries sunken into upper part of ovary. Disc absent.
Androecium Stamens five (median adaxial absent), in a single whorl. Filaments free, adnate at base to inner tepals. Anthers basifixed, non-versatile, tetrasporangiate, introrse, longicidal (dehiscing by longitudinal slits); connective somewhat prolonged at apex. Tapetum secretory? Staminodium usually absent.
Pollen grains Microsporogenesis successive? Pollen grains inaperturate, shed as monads, ?-cellular at dispersal. Exine intectate, psilate.
Gynoecium Pistil composed of three connate carpels. Ovary inferior, trilocular, prolonged at apex. Style single, simple, elongate, widened at apex. Stigma trilobate, zygomorphic, dorsiventrally flattened, with secretory tissue, ‘viscidium’, at base of adaxial/ventral side and with entire, lobate or fimbriate stigmatic lobes, non-papillate, Wet type. Pistillodium absent.
Ovules Placentation axile. Ovules c. 30 to more than 50 per carpel, anatropous, bitegmic, crassinucellar. Micropyle ?-stomal. Outer integument 14 to 16 cell layers thick. Inner integument approx. four cell layers thick. ‘Micropylar collar’ developed? Parietal cell? Megagametophyte monosporous, Polygonum type? Endosperm development ab initio nuclear. Endosperm haustoria? Embryogenesis?
Fruit A loculicidal capsule with lower part of ovary persistent.
Seeds Aril lacerate, consisting of three or four coarse hair-like outgrowths. Testa vascularized, with operculum opposite radicula. Seed coat testal. Exotesta and adjacent two inner layers lignified; exotesta with phytomelan on cell walls. Endotesta silicified?, consisting of radially elongate sclereids. Tegmen crushed? Perisperm thin, slightly developed. Endosperm copious, with starch grains. Embryo cylindrical, well differentiated, Trillium type, chlorophyll? Cotyledon one, not assimilating? Cotyledon hyperphyll? Hypocotyl internode? Coleoptile? Collar? Germination?
Cytology n = 9
DNA
Phytochemistry Virtually unknown. Tannins present. Cyanogenic compounds not found.
Use Unknown.
Systematics Orchidantha (16; southernmost China, Hainan, Southeast Asia, the Malay Peninsula, Borneo, the Philippines, islands in western Pacific).
Orchidantha is probably sister to Strelitziaceae (Cron & al. 2012).
MARANTACEAE R. Br. |
( Back to Cannales ) |
Marantales R. Br. in C. F. P. von Martius, Consp. Regn. Veg.: 10. Sep-Oct 1835 [‘Marantaceae’]
Genera/species 25–27/320–340?
Distribution Pantropical except Australia, with their largest diversity in tropical America.
Fossils Unknown.
Habit Bisexual, perennial (sometimes giant) herbs (rarely climbing; sometimes with woody rhizome). Rhizome rich in starch.
Vegetative anatomy Roots usually fibrous (rarely with nodules). Phellogen absent. Secondary lateral growth absent. Vessels present in roots (sometimes also in stems). Vessel elements with scalariform and/or simple perforation plates; lateral pits? Imperforate tracheary xylem elements tracheids. Wood rays usually absent. Axial parenchyma? Sieve tube plastids P2cs type, with cuneate protein crystals and starch grains, without protein filaments. Nodes? Mucilage cells and secretory cells rare. Mucilage canals usually absent (present in Thalia). Silica bodies with irregular outgrowths, conical or druse-like, in stegmata or thin-walled cells. Calciumoxalate as single, rhomboidal, prismatic or rod-shaped crystals (raphides absent). Starch grains usually elliptic and somewhat excentric (rarely flat or angular), single or aggregated.
Trichomes Hairs usually absent (sometimes unicellular).
Leaves Alternate (distichous, sometimes in right angles to stem), simple, entire, with supervolute ptyxis (laterally rolled-up in bud?), usually differentiated into often long pseudopetiole (often absent) and asymmetrical pseudolamina, with a pulvinus (with oblique cells) in transition zone between pseudopetiole and pseudolamina. Stipules absent; leaf sheath closed, without ligule. Venation pinnate-parallelodromous, with distinct mid-vein and closely spaced parallel arcuate lateral veins converging and fused at leaf margin and connected by transverse tertiary veins. Stomata tetracytic or paracytic (sometimes tricytic, anomocytic or polycytic); guard cells with inner and outer borders non-uniform. Cuticular wax crystalloids usually absent (sometimes non-orientated). Epidermis without silica bodies. Mesophyll without mucilaginous idioblasts; mesophyll cells with calciumoxalate as druses or single prismatic crystals (raphides absent). Leaf margin entire. Extrafloral nectaries present at apex of leaf sheath in some species of Ctenanthe and Stromanthe.
Inflorescence Usually terminal (sometimes axillary), simple or branched; each partial inflorescence – usually a spike- or head-like thyrse – consisting of groups (subtended by bracts) of one or several (rarely numerous) usually two-flowered (in Monotagma and Monophrynium one-flowered) cymules; each cymule with a basal adaxial prophyll (sometimes also with a scale-like abaxial interphyll and often one or two adaxial bracteoles); each cymule an enantiomerous pseudanthium with two asymmetrical flowers in each pair being mirror images of each other. Floral prophyll (bracteole) absent or lateral: on right side of first flower and on left side of second flower in each pair, sometimes with apical extrafloral nectaries.
Flowers Asymmetrical, often small, resupinate. Epigyny. Tepals 3+3; outer tepals sepaloid, free; inner tepals petaloid, connate at base into a tube, adnate to androecium and style; median outer perianth member adaxial; median inner tepal often dome- or hood-shaped, and larger than the rest. Septal/gynoecial nectaries sunken at ovary apex. Disc absent.
Androecium Outer staminal whorl consisting of one or two lateral sometimes petaloid staminodia, or absent; median adaxial outer stamen absent. Inner staminal whorl consisting of one fertile (adaxial inner) stamen, with a single lateral theca (two microsporangia) and often with a lateral petaloid outgrowth (transformed sterile theca?), and two petaloid staminodia: one sensitive dome- or hood-shaped staminodium, staminodium cucullatum (usually lobed and with lateral appendages), enclosing style and stigma prior to pollination, and a second staminodium, staminodium callosum, (hard, fleshy and warty on adaxial side). Filaments of outer staminodia connate into a tube, adnate at base to tepals and style; filament of inner stamen and staminodia connate at base into a tube. Anther dorsifixed?, non-versatile, disporangiate, introrse, longicidal (dehiscing by longitudinal slits). Tapetum amoeboid-periplasmodial? (or secretory?), with ?-nucleate cells.
Pollen grains Microsporogenesis successive. Pollen grains inaperturate, shed as monads, bicellular at dispersal. Exine very thin or absent; pollen wall consisting of intine. Pollen surface psilate.
Gynoecium Pistil composed of three connate carpels, two of which often as sterile pistillodia. Ovary inferior, trilocular, with two locules often compressed and sterile. Style single, subulate, adnate at base to perianth-staminal tube; a flat adaxial subapical surface delimited on abaxial side by gland-like secretory swelling; style held under tension by staminodium cucullatum prior to pollination. Pollen receptive surface (‘stigma’) present on inner side of an infundibuliform subapical cavity, non-papillate, Wet type. Pistillodia present or absent. Secondary pollen presentation (pollen grains deposited from developing theca on adaxial secretory surface prior to anthesis).
Ovules Placentation basal to axile. Ovule one per carpel, ab initio anatropous, subsequently amphitropous (anacampylotropous), bitegmic, crassinucellar. Micropyle usually endostomal (in Phrynium bistomal). Outer integument six to eight (to twelve) cell layers thick. Inner integument ? (one or two?) cell layers thick. A cylindrical ridge-shaped thickening, ‘micropylar collar’ (‘Mikropylarkragen’), formed from outer integument surrounding micropyle and penetrating micropylar part of megasporangium. Operculum formed by periclinal cell divisions outside of ‘collar’. Parietal cell formed from archesporial cell. Chalazal (vascular) tissue growing into megasporangium. Nucellar cap (sometimes approx. two cell layer thick) formed from palisade-like megasporangial epidermis. Lateral epidermal cells dividing periclinally. Megagametophyte monosporous, Polygonum type. Antipodal cells sometimes proliferating. Endosperm development ab initio nuclear. Endosperm haustoria? Embryogenesis chenopodiad.
Fruit Usually a loculicidal capsule (in Sarcophrynium and Thaumatococcus baccate; in Thalia nutlike).
Seeds Aril formed from outer epidermis of outer integument near micropyle, or absent. Seed coat testal. Exotesta consisting of fibriform elongate cells? and with phytomelan layer on epidermal cell walls. Mesotesta tanniniferous. Endotestal cells with silica. Operculum formed from endotestal epidermis and combined with ‘micropylar collar’ (cf. Canna). Tegmen crushed, sometimes with thin elongate sclereids. Perisperm hard, copious, with thick-walled cells containing starch grains. Chalazal tissue, chalazosperm, often well developed, with a branched or simple perisperm canal formed in megasporangium by degradation of chalazal tissue. Endosperm rudimentary or absent. Embryo long, curved, without chlorophyll, Trillium type. Cotyledon one. Cotyledon hyperphyll compact, not assimilating. Hypocotyl internode absent. Mesocotyl sometimes present; collar inserted at right angles to cotyledon. Coleoptile present. Germination?
Cytology n = 9–13 (chromosome numbers uncertain!)
DNA Mitochondrial coxI intron present in several genera.
Phytochemistry Flavonols (kaempferol, quercetin), flavonol sulphates, flavone-C-glycosides, luteolin, anthocyanins, deoxyanthocyanins (apigenin), cyanidin, delphinidin, pelargonidin, tannins, and cyanogenic compounds present. Thaumatins (high-molecular proteins) present in aril (1.600 times sweeter than saccharose). Ellagic acid, chelidonic acid, and saponins not found.
Use Ornamental plants, starch sources (West Indian arrowroot from Maranta arundinacea), vegetables and thatching (Calathea), basketry (Ischnosiphon).
Systematics Sarcophrynium (6; S. bisubulatum, S. brachystachyum, S. congolense, S. prionogonium, S. schweinfurthianum, S. villosum; tropical Africa), Megaphrynium (5; M. distans, M. gabonense, M. macrostachyum, M. trichogynum, M. velutinum; tropical West and Central Africa), Thaumatococcus (1; T. daniellii; tropical West Africa), Hypselodelphys (8; tropical West and Central Africa), Trachyphrynium (1; T. braunianum; tropical West Africa); Haumania (5; H. danckelmaniana, H. leonardiana, H. liebrechtsiana: tropical West and Central Africa; H. microphylla: New Guinea; H. walkeri: Japan); Monotagma (37; tropical South America), Pleiostachya (3; P. leiostachya, P. pittieri, P. pruinosa; southern Mexico, Central America, Colombia, Ecuador), Ischnosiphon (c 35; tropical America),’Calathea’ pro parte (45–50?; tropical America); Thalia (6; T. dealbata, T. densibracteata, T. geniculata, T. multiflora, T. pavonii, T. petersiana; southeastern United States to northern Argentina, one species, T. geniculata, also in Central America, the West Indies and tropical Africa), Donax (1; D. canniformis; Southeast Asia, Malesia to Vanuatu), Schumannianthus dichotomus (India to Southeast Asia and the Philippines), Phrynium (35–40; India to southern China and Southeast Asia, New Guinea, Vanuatu); Halopegia (3; H. azurea: Congo; H. blumei: Indochina, Java; H. perrieri: Madagascar), Schumannianthus virgatus (southern India, Sri Lanka), Hylaeanthe (5; H. hexantha, H. hofmannii, H. panamensis, H. polystachya, H. unilateralis; Central America, tropical South America), Maranta (40–45; southern Mexico, Central America, the West Indies, tropical South America), Myrosma (1; M. cannifolia; tropical South America), Saranthe (10; Brazil), Ctenanthe (c 10; Central America, tropical South America), Stromanthe (c 20; southern Mexico, Central America, tropical South America); Monophyllanthe (2; M. araracuarensis, M. oligophylla; northern South America), Marantochloa (18; tropical Africa, Madagascar, the Comoros), Stachyphrynium (8; India and Sri Lanka to Tibet, Yunnan, southeastern China, Andaman Islands, Southeast Asia, West Malesia to Sulawesi), Afrocalathea (1; A. rhizantha; Central Africa).
Marantaceae are sister-group to Canna (Cannaceae). The Sarcophrynium clade is sister to the remaining Marantaceae (Prince & Kress 2006).
The trend towards secondary pollen presentation may be a synapomorphy of Marantaceae and Canna. In Marantaceae it is a type of explosive mechanism. The pollen grains are deposited from the developing theca into a cavity on the adaxial side near the apex of the style, which is curved and held under tension by the staminodium cucullatum. The secretions from the adjacent glands make the pollen grains stick to the stylar surface. The appendages of the staminodium are touched by the nectar-searching pollinator, and the style triggered and momentarily (within a few thousands of a second) recurved releasing the deposited pollen. Simultaneously, pollen grains from a different flower may be transferred from the pollinator to the receptive surface.
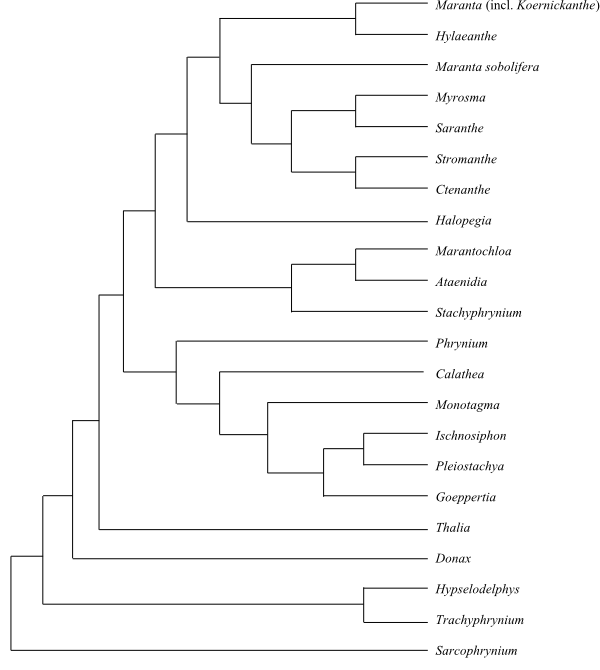 |
Cladogram of Marantaceae based on DNA sequence data and morphology (Andersson & Chase 2001; Borchsenius & al. 2012). |
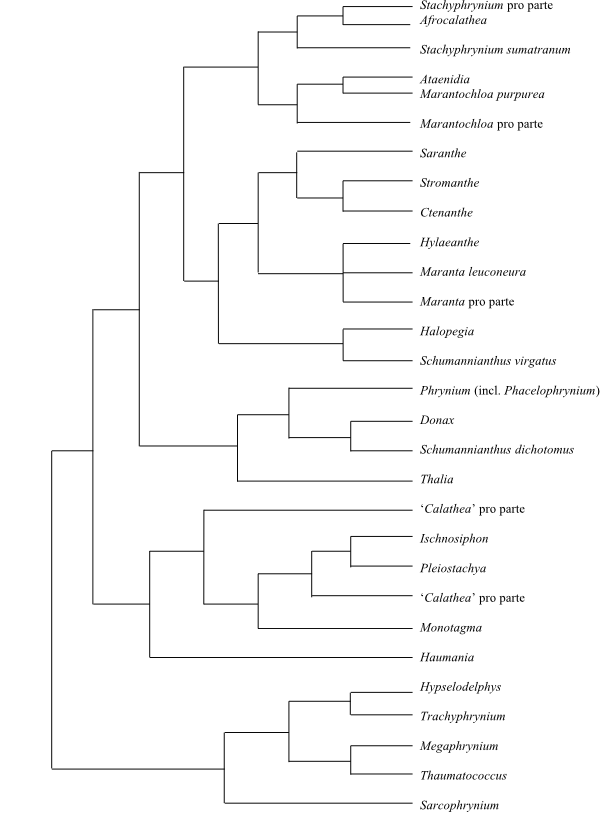 |
Cladogram (one of 48 shortest maximum parsimony trees) of Marantaceae based on DNA sequence data (Prince & Kress 2006). |
MUSACEAE Juss. |
( Back to Cannales ) |
Musales Juss. ex Bercht. et J. Presl, Přir. Rostlin: 269. Jan-Apr 1820 [‘Musae’]
Genera/species 2/40–45
Distribution Tropical Africa, Madagascar, tropical Asia to Queensland, islands in western Pacific.
Fossils Uncertain. Fossil remains from the Eocene of Oregon have been assigned to Ensete.
Habit Usually monoecious or andromonoecious, with male flowers at apex and usually female flowers (rarely bisexual flowers) at base of inflorescence, perennial giant herbs with aerial pseudostem consisting of leaf sheaths (species in Ensete hapaxanthic, monocarpous). Rhizome rich in starch, with short and thick branches. Axillary buds leaf-opposite or absent.
Vegetative anatomy Root cortex sometimes with periderm. Root medulla with scattered wide vessels and phloem strands. Endodermis present in rhizome. Phellogen absent. Secondary lateral growth absent. Vessels present in roots and rhizome. Vessel elements with scalariform and/or simple perforation plates; lateral pits? Imperforate tracheary xylem elements tracheids. Wood rays absent. Axial parenchyma? Sieve tube plastids P2cfs type, with cuneate protein crystals, starch grains and protein filaments. Articulated laticifers with mucilage (becoming dark when oxidized) numerous in Musa in association with vascular bundles (not in roots). Silica bodies boat-shaped, in association with vascular bundles (not so in roots). Idioblasts (raphide sacs) with calciumoxalate raphides. Starch grains simple, concentric.
Trichomes Hairs absent from vegetative parts.
Leaves Alternate (spiral), simple, entire, with supervolute ptyxis (laterally rolled-up in bud?), usually differentiated into pseudopetiole and pseudolamina. Stipules absent; leaf sheath well developed, without distinct ligule. Pseudopetiole with a single row of abaxial air canals. Venation pinnate-parallelodromous; mid-vein distinct; secondary veins closely spaced, parallel, arising in almost right angles from mid-vein, converging at leaf margin and connected by transversal tertiary veins. Stomata usually paracytic (sometimes tetracytic). Cuticular wax crystalloids as longitudinally aggregated rodlets (Strelitzia type), chemically dominated by wax esters or alkanes. Epidermis without silica bodies. Mesophyll with mucilaginous idioblasts containing calciumoxalate as raphides and single rhomboidal crystals. Leaf margin entire.
Inflorescence Terminal, compound spike-like thyrse consisting of lateral cincinni in axils of large, caducous (in Musa) or persistent (in Ensete), spatha-like and boat-shaped bracts. Thyrse with basal female (rarely bisexual) flowers and apical male flowers. Prophylls lateral. Floral bracts and prophyll (bracteole) absent. Extrafloral nectaries sometimes present.
Flowers Zygomorphic, large. Epigyny. Tepals 3+3, petaloid, pseudomonocyclic; median outer tepal abaxial (anterior); three outer and two lateral inner tepals connate into a tube or spatha; median inner tepal adaxial (posterior), free, small and dome-shaped (sometimes deeply concave), directed downwards. Septal/gynoecial nectaries present in upper part of ovary. Disc absent.
Androecium Stamens five, with inner median adaxial (posterior) stamen small or staminodial (in Musa), or stamens six (in Ensete), antetepalous. Filaments filiform, free from each other and from tepals. Anthers basifixed, non-versatile, tetrasporangiate, introrse, longicidal (dehiscing by longitudinal slits); connective occasionally somewhat prolonged. Tapetum secretory, with binucleate cells. Staminodium one (opposite free stamen) or absent; female flowers with staminodia.
Pollen grains Microsporogenesis successive. Pollen grains inaperturate (omniaperturate), shed as monads, bicellular at dispersal. Exine very thin to almost absent; pollen surface psilate (in Musa) or gemmate to verrucate (in Ensete). Intine thick.
Gynoecium Pistil composed of three connate carpels; median carpel abaxial (anterior). Ovary inferior, trilocular, in Musa with mucilage and hairs on placentae (sometimes also on funicle). Style single, simple, filiform, often widened at apex. Stigma capitate, papillate, Wet type. Male flowers with pistillodium.
Ovules Placentation axile. Ovules approx. ten to more than 50 per carpel, anatropous, bitegmic, crassinucellar. Micropyle sometimes exostomal. Outer integument six to nine cell layers thick. Inner integument two or three cell layers thick. A cylindrical ridge-shaped thickening, ‘micropylar collar’ or ‘Mikropylarkragen’, formed from outer integument surrounding micropyle. Hypostase present. Parietal cell not formed (parietal tissue absent). Megagametophyte monosporous, Polygonum type. Endosperm development ab initio nuclear. Endosperm haustoria? Embryogenesis?
Fruit A berry-like capsule with leathery pericarp, indehiscent or partially dehiscing longitudinally. Seeds surrounded by a fleshy starchy pulp formed from placental hairs.
Seeds Funicular aril reduced, consisting of simple hairs embedded in mucilage, or absent. Seed with chalazal space. Exotestal cells silicified, forming dense sclerenchyma. Mesotesta massive, 20–25 cell layers thick, sclerotic. Endotestal cells silicified? Tegmen two cell layers thick, with elongate cells. Operculum situated above micropylar part and combined with ‘micropylar collar’. Perisperm one cell layer thick, with lipids (starch absent). Endosperm copious, starchy (with small chalazal space?). Embryo straight (in Musa) or somewhat curved (in Ensete), fungiform, Trillium type, chlorophyll? Cotyledon one. Cotyledon hyperphyll compact, not assimilating. Hypocotyl internode relatively well developed. Mesocotyl absent. Coleoptile present. Double micropylar collar well developed (at right angles to cotyledon). Germination phanerocotylar.
Cytology n = 7, 9–11 (Musa); n = 9, 10 (Ensete)
DNA Mitochondrial coxI intron present.
Phytochemistry Flavonols (kaempferol, quercetin), cyanidin, pelargonidin, malvidin and malvidin glycoside, anthoxanthins, deoxyanthocyanins, tannins, proanthocyanidins (prodelphinidins), indole alkaloids, cyanogenic glycosides, and phenylphenalenones (perinaphthenones) present. Ellagic acid and saponins not found.
Use Ornamental plants, fruits and vegetables, textiles, carpets and rope (abaca, manila-hemp from Musa textilis), beer, etc.
Systematics Musa (35–40; eastern and southern India, Sri Lanka, southernmost China, Southeast Asia, Malesia to New Guinea and northeastern Queensland, islands in western Pacific), Ensete (6–8; E. gilletii, E. glaucum, E. homblei, E. perrieri, E. superbum, E. ventricosum, E. wilsonii; tropical and subtropical regions in Africa, Madagascar and Asia to New Guinea; incl. Musella).
Possibly, Musaceae are sister to the remaining Cannales, but the support is low.
Musella is nested within Ensete in analyses by Liu & al. (2010).
STRELITZIACEAE (K. Schum.) Hutch. |
( Back to Cannales ) |
Genera/species 3/7
Distribution South Africa, Madagascar, tropical South America.
Fossils Uncertain. The fossil genus Cyclanthodendron (including fruits with the name Tricoccites) from the Eocene of India? has sometimes been attributed to Cyclanthaceae, but possibly is more allied to Strelitziaceae.
Habit Bisexual, perennial (often giant) herbs (aerial stem in Phenakospermum, Ravenala and Strelitzia nicolai lignified). Rhizome rich in starch, with short and thick branches. Dichotomous branching present in Strelitzia.
Vegetative anatomy Root medulla with uniformly scattered wide xylem and phloem strands. Aerial stems with superficial mechanical layer and with vascular bundles surrounded by dense fibre envelopes. Phellogen absent? Central vascular cylinder not delimited by endodermis. Secondary lateral growth absent. Vessels present in roots and stem (in Ravenala only in roots). Vessel elements usually with scalariform (in Phenakospermum also simple) perforation plates; lateral pits? Imperforate tracheary xylem elements tracheids. Wood rays? Axial parenchyma? Sieve tube plastids P2cs type, with cuneate protein crystals and starch grains, without protein filaments. Nodes? Tannin frequent in normal cells. Silica bodies, druse-shaped or spherical, present in hypodermal cells in association with vascular bundles (not in roots), or spherical in epidermal stegmata. Idioblasts (raphide sacs) with calciumoxalate raphides. Starch grains simple, usually spherical (in Ravenala flattened).
Trichomes Hairs absent.
Leaves Alternate (distichous), simple, entire, with supervolute ptyxis (laterally rolled-up in bud?), usually differentiated into pseudopetiole and pseudolamina. Stipules absent; leaf sheath well developed, without distinct ligule. Pseudopetiole long, with several adaxial and abaxial arcs of air canals. Venation pinnate-parallelodromous; midvein distinct; secondary veins closely spaced, parallel, converging at leaf margin and connected by transversal tertiary veins. Stomata polycytic, tetracytic or hexacytic, with sometimes oblique cell divisions and often sunken guard cells. Cuticular wax crystalloids as longitudinally aggregated terete rodlets (Strelitzia type), chemically dominated by wax esters, usually concentrated around stomata. Epidermis without silica bodies. Mesophyll with mucilaginous idioblasts containing calciumoxalate raphides. Leaf margin entire.
Inflorescence Terminal (Phenakospermum) or lateral (Ravenala, Strelitzia), compound spike-like thyrse consisting of one or several distichous cincinni in axils of large stout persistent spatha-like bracts. Bracts and floral prophylls (bracteoles) boat-shaped to lanceolate.
Flowers Zygomorphic, large. Epigyny. Tepals 3+3; median outer adaxial (posterior); two lateral inner tepals larger than median adaxial (cucullate) inner tepal; outer tepals free; inner tepals connate at base; in Strelitzia two lateral abaxial inner tepals connate into a sagittate structure surrounding stamens and style. Septal/gynoecial nectaries deeply sunken into upper part of ovary. Disc absent.
Androecium Stamens 3+2 (median inner adaxial stamen staminodial; Phenakospermum, Strelitzia) or 3+3 (Ravenala), antetepalous (at least in Strelitzia). Filaments free from each other and from tepals. Anthers basifixed, non-versatile, tetrasporangiate, introrse, longicidal (dehiscing by longitudinal slits); connective in Strelitzia somewhat prolonged; anthers in Strelitzia with filiform structures inside; anthers in Phenakospermum up to 7 cm long, present in perianth keel and first exposed at pollination (by bats). Tapetum secretory, with up to 32-ploid cells. Staminodium one (adjacent to adaxial inner tepal) or absent.
Pollen grains Microsporogenesis successive. Pollen grains inaperturate (omniaperturate), shed as monads, unicellular at dispersal (vegetative cell degenerating). Exine very thin to almost absent; pollen surface psilate? Intine thick. Pollen wall in Phenakospermum discontinuous.
Gynoecium Pistil formed by three connate carpels. Ovary inferior, trilocular, prolonged at apex. Style single, simple, filiform, often widened at apex. Stigma long, turbinate, or trilobate, papillate, Wet type. Pistillodium absent.
Ovules Placentation axile. Ovules c. 20 to more than 50 per carpel, anatropous, bitegmic, crassinucellar. Micropyle bistomal. Outer integument ? cell layers thick. Inner integument ? cell layers thick. Cylindrical ridge-shaped thickening, ’micropylar collar’ (‘Mikropylarkragen’), absent. Hypostase absent. Parietal cell absent or formed from archesporial cell, not forming distinct parietal layer. Megagametophyte monosporous, Polygonum type. Endosperm development ab initio nuclear. Endosperm haustoria? Embryogenesis?
Fruit A lignified loculicidal capsule.
Seeds Aril exostomal, fimbriate to lobate, sometimes brightly coloured, formed partly from outer integument near micropyle, partly from funicle opposite micropyle. Seed coat testal. Testa with compact outer cuticle. Inner testal epidermis consisting of large cells with inner tangential walls strongly thickened (U-cell layer). Tegmen consisting of cuticle (remaining layers crushed). Operculum rudimentary or absent. Perisperm one-layered, reduced, with lipids (starch absent), or absent. Endosperm copious, starchy, outer cell layers with aleurone. Embryo straight or curved, well differentiated, Trillium type, chlorophyll? Cotyledon one, massive. Cotyledon hyperphyll compact, not assimilating. Hypocotyl internode and mesocotyl absent. Coleoptile present. Collar? Radicula well developed, persistent. Germination?
Cytology n = 7, 9, 11
DNA
Phytochemistry Flavonols (kaempferol, quercetin), cyanidin, tannins, phenylphenalenones, and oxypipe colanic acid (in Strelitzia) present. Flavonols (kaempferol, quercetin) and cyanidin absent from Ravenala. Ellagic acid and cyanogenic compounds not found.
Use Ornamental plants.
Systematics Ravenala (1; R. madagascariensis; Madagascar), Phenakospermum (1; P. guyanense; eastern tropical South America), Strelitzia (5; S. alba, S. caudata, S. nicolai, S. reginae, S. juncea; temperate and subtropical regions of southern Africa).
Strelitziaceae are probably sister to Orchidantha(Lowiaceae) (Cron & al. 2012).
Prince (2010) presents a cladogram of Cannales, including the following topology of Strelitziaceae: [Ravenala+[Phenakospermum+Strelitzia]]. On the other hand, Cron & al. (2012) were not able to unequivocally define the sister-group relationship in Strelitziaceae. Ravenala was sister to [Phenakospermum+Strelitzia] or Strelitzia was sister-group to [Phenakospermum+ Ravenala]. Within Strelitzia, S. nicolai is sister to the remaining species.
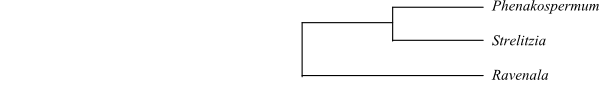 |
Cladogram of Strelitziaceae based on DNA sequence data (Prince 2010; Cron & al. 2012) |
ZINGIBERACEAE Martinov |
( Back to Cannales ) |
Amomaceae J. St.-Hil., Expos. Fam. Nat. 1: 154. Jan-Aug 1805, nom. illeg.; Alpiniaceae Link, Enum. Hort. Berol. Alt. 1: 2. 16 Mar-30 Jun 1821; Curcumaceae Dumort., Anal. Fam. Plant.: 56. 1829; Amomales Lindl., Key Bot.: 69. 15-30 Sep 1835
Genera/species c 62/1.470–>1.510
Distribution Tropical and southern Africa, Madagascar, islands in the Indian Ocean, South, East and Southeast Asia, Malesia to New Guinea, tropical Australia (especially Queensland), islands in the Pacific; some species of Renealmia in tropical America (Central America, the West Indies, northern and central South America).
Fossils Fossil fruits, seeds and leaves, Zingiberopsis, of plausible Zingiberaceae are known from Campanian and early Palaeogene layers in North America.
Habit Bisexual, perennial (sometimes giant) herbs, often with a pseudostem consisting of superimposed leaf sheaths. Sometimes with stilt roots. Aromatic. Rhizome and roots usually rich in starch (root cells sometimes with starch nodules).
Vegetative anatomy Phellogen absent. Stem cortex and central tissue with each vascular bundle surrounded by a dense fibre envelope; cortex and central tissue separated by a cylinder of fibres. Secondary lateral growth absent. Vessels present in roots (rarely also in stem). Vessel elements usually with scalariform (sometimes also simple) perforation plates; lateral pits? Imperforate tracheary xylem elements tracheids. Wood rays absent. Axial parenchyma? Sieve element nucleus with non-dispersive crystalline protein bodies. Sieve tube plastids P2cs type, with cuneate protein crystals and starch grains, without protein filaments. Nodes? Idioblasts with suberised cell walls and containing abundant aromatic oils and resins. Stegmata-shaped cells with single large silica bodies sometimes present (in Alpinioideae in connection with vascular bundles). Calciumoxalate as rhomboidal, prismatic or tubular crystals (raphides usually absent). Crystal sand present at least in Roscoea. Starch grains usually simple, often flat (sometimes irregular), usually excentric (sometimes compound; rarely spherical or elliptic).
Trichomes Hairs usually absent, when present usually unicellular (on rhizome in Curcuma bicellular; rarely with sunken base; rarely multicellular, furcate to stellate or uniseriate).
Leaves Alternate (distichous), simple, entire, often differentiated into pseudopetiole and pseudolamina, with supervolute ptyxis (laterally rolled-up in bud). Stipules absent; leaf sheath usually open (sometimes closed), usually with a ligule at distal end. Pulvinus usually absent (present in, e.g., Zingiber). Pseudopetiole and mid-vein with cylinder of aerenchyma strands. Vascular bundles abaxial. Venation pinnate-parallelodromous; mid-vein distinct; secondary veins closely spaced, parallel, connected by transverse tertiary veins. Stomata usually tetracytic (sometimes hexacytic or paracytic); guard cells with inner and outer borders non-uniform. Cuticular wax crystalloids usually absent (sometimes present as platelets or rodlets). Epidermis with spherical silica bodies or silica sand. Mesophyll with spherical idioblasts containing ethereal oils and calciumoxalate as druses or prismatic crystals (raphides usually absent). Leaf margin entire.
Inflorescence Flowers on foliar shoots or on special non-photosynthesizing shoots, in terminal usually head- or cone-like (rarely raceme- och spike-like) often branched thyrse consisting of few- to many-flowered (sometimes one-flowered) cincinni subtended by spirally arranged imbricate bracts (bracts rarely absent). Floral prophyll (bracteole) sometimes lateral. Extrafloral nectaries frequent.
Flowers Zygomorphic, often large. Epigyny. Tepals 3+3; outer tepals usually sepaloid, partially connate into a cupulate or asymmetrical spatha-like structure with a lateral longitudinal slit; median inner tepal adaxial, dome-shaped and usually larger than lateral inner tepals; inner tepals petaloid, connate below into a tube (outer and inner tepals in species of Cyphostigma and Elettaria together forming an outgrowth on top of ovary). Gynoecial/septal epigynous nectaries two, inserted at ovary apex; one or three glandular nectaries present at stylar base and staminal bases (sometimes also on staminodia, androecial nectaries). Disc absent.
Androecium Stamen single (median inner adaxial stamen fertile; median outer abaxial stamen absent). Filament of fertile stamen short or long (sometimes absent). Anther dorsifixed, sometimes versatile, tetrasporangiate, introrse, usually longicidal (dehiscing by longitudinal slits; rarely poricidal, dehiscing by pores); connective at apex usually prolonged into a pectinate structure of varying shape (in Kaempferia petaloid); placentoid present. Tapetum usually secretory (sometimes amoeboid-periplasmodial), with uninucleate or binucleate cells. Staminodia two or four; two lateral inner staminodia connate into a petaloid labellum; two lateral outer staminodia petaloid or small and reduced (in some genera absent), inserted on both sides of fertile stamen or adnate to labellum; inner tepals adnate to staminodia in Globba and Cautleya; inner tepals and labellum free from each other in Vanoverbergia, Riedelia, Aframomum, Hornstedtia, Zingiber, Amomum, and Cautleya; filament and labellum fused into a tube in Geocharis, Pommereschea, Etlingera, Aulotandra, Stadiochilus, and Globba; lateral staminodia adnate to filament of functioning stamen in Mantisia; staminodia and labellum fused in Siphonochilus.
Pollen grains Microsporogenesis successive. Pollen grains usually inaperturate (sometimes monosulcate or 5–16-pantoporate), shed as monads, bicellular at dispersal. Exine tectate (tectum thin; exine sometimes intectate), with ? infratectum, verrucate, echinate, scabrate, rugulate, striate or smooth, or sometimes absent.
Gynoecium Pistil composed of three connate carpels; median carpel abaxial. Ovary inferior, ab initio trilocular, finally incompletely or entirely trilocular or unilocular. Style single, filiform, hollow, situated in a groove in filament of fertile stamen and between the two anther halves (thecae). Stigma infundibuliform, papillate or non-papillate, Wet type, present above fertile anther. Pistillodium absent.
Ovules Placentation axile (when ovary trilocular); in ‘Globbeae’ parietal with unilocular ovary; in some genera of Zingibereae basal through reduction of septa. Ovules (one to) four to c. 50 per carpel, anatropous or orthotropous, bitegmic, crassinucellar. Micropyle endostomal. Outer integument (five to) seven to 13 cell layers thick. Inner integument ? cell layers thick. A cylindrical ridge-shaped thickening, ‘micropylar collar’ (‘Mikropylarkragen’), formed from outer integument surrounding micropyle. Hypostase present in many species. Epistase usually present. Nucellar cap sometimes present. Parietal cell formed from archesporial cell. Lateral epidermal cells dividing periclinally. Megagametophyte monosporous, Polygonum type. Endosperm development ab initio helobial; chalazal chamber early degenerating. Endosperm haustoria? Embryogenesis asterad.
Fruit Usually a loculicidal, dry or fleshy capsule (sometimes irregularly dehiscing or indehiscent; in Aframomum a berry; in, e.g., Etlingera a fleshy syncarp).
Seeds Aril usually present. Seed coat testal. Exotesta consisting of fibriform elongate cells and with phytomelan layer on epidermal cell walls. Endotestal cells with silica, usually sclerotic and U-shaped in cross-section (in Globba, Hedychium and their closest relatives thin, not U-shaped). Ethereal oils present in testa. Operculum (formed from inner testal epidermis) present adjacent to radicula (and combined with ‘micropylar collar’). Perisperm well developed, copious, rich in compound starch grains. Chalazosperm absent. Endosperm usually sparse, with aleurone (sometimes with starch grains). Embryo straight (in Elettaria large), without chlorophyll, without lipid bodies, Trillium type. Cotyledon one. Hypocotyl internode short to long. Mesocotyl absent. Cotyledon hyperphyll compact, not assimilating. Coleoptile present or absent. Collar indistinct. Germination cryptocotylar?
Cytology n = 8–14, 17, 21, 25 – Polyploidy and aneuploidy frequently occurring.
DNA Two different indel mutations (insertions), each one comprising one base-pair in plastid 18SrDNA, present in all investigated Zingiberaceae, but not found in other taxa (Soltis & al. 1997). Mitochondrial intron coxII.i3 lost (Kaempferia). Mitochondrial coxI intron present in several genera (including Siphonochilus).
Phytochemistry O-methylated flavonols (kaempferol, myricetin, isorhamnetin, syringetin) flavanones, chalcones, ethereal oils consisting of mono-, di-, tri- and sesquiterpenes, condensed tannins (from proanthocyanidins), proanthocyanidins (prodelphinidin, procyanidin), phenylpropanoids and related curcumins (diarylheptanoids), caffeic acid, quinines, glycosides (e.g. 3-glycoside, 3-rutenoside), aromatic acids and ketones, and shikimic acid- or polyacetate-derived arthroquinones present. Dihydroflavones, alkaloids, steroidal saponins, and cyanogenic compounds rare. Flavone-C-glycosides and ellagic acid not found.
Use Ornamental plants, starch sources (East Indian arrowroot from Curcuma), spices (Aframomum melegueta, Amomum, Boesenbergia, Curcuma, Elettaria cardamomum, Kaempferia, Zingiber officinale, etc.), medicinal plants, dyeing sources (Curcuma), perfumes (Alpinia galanga, A. officinale, Curcuma zedoaria, Hedychium).
Systematics Zingiberaceae are sister to Costaceae. The African-Malagasy Siphonochilus is sister to the remaining Zingiberaceae.
Siphonochiloideae W. J. Kress in W. J. Kress, L. M. Prince et K. J. Williams in Amer. J. Bot. 89: 1691. 2002
1/11. Siphonochilus (11; tropical and southern Africa, Madagascar). – Rhizome fleshy, vertical. Distichy of leaves perpendicular to rhizome. Inflorescence a raceme. Floral prophylls (bracteoles) absent. Lateral staminodia petaloid, adnate to labellum. Labellum not connate to filament. Anther crest petaloid. Ovary trilocular (sometimes inclompletely so). Placentation axile. Fruit fleshy. n = ?
[Tamijia+[Alpinioideae+Zingiberoideae]]
Tamijioideae W. J. Kress in W. J. Kress, L. M. Prince et K. J. Williams in Amer. J. Bot. 89: 1691. 2002
1/1. Tamijia (1; T. flagellaris; northwestern Borneo). – Rhizome fibrous. Distichy of leaves perpendicular to rhizome. Lateral staminodia petaloid, adnate to labellum. Labellum not connate to filament. Anther crest petaloid. Ovary unilocular. Placentation parietal. Fruit? n = ? – Tamijia is sister to the main clade [Alpinioideae+Zingiberoideae].
[Alpinioideae+Zingiberoideae]
Labellum formed exclusively of the two staminodia in the inner androecial whorl. Placentation sometimes free central. Endosperm without starch.
Alpinioideae (Blume) Hassk., Cat. Hort. Bot. Bogor.: 51. Oct 1844 [‘Alpiniae’]
c 27/850–>870. Riedelieae in W. J. Kress, L. M. Prince et K. J. Williams in Amer. J. Bot. 89: 1691. 2002. Siliquamomum (1; S. tonkinense; Vietnam), Siamanthus (1; S. siliquosus; Thailand), Burbidgea (5; B. longiflora, B. nitida, B. pauciflora, B. schizocheila, B. stenantha; Borneo), Pleuranthodium (23; New Guinea, one species, P. peekelii, in the Bismarck Archipelago and one species, P. racemigerum, in eastern Queensland), Riedelia (c 75; East Malesia to New Guinea and Solomon Islands). – Alpinieae (Blume) Orb., Dict. Univ. Hist. Nat. 1: 299. 18 Dec 1841. ’Alpinia’ (230–240; tropical and subtropical Asia to islands in the Pacific; non-monophyletic), Aframomum (50–55; tropical Africa, Madagascar, the Mascarene Islands, the Seychelles), Elettaria (1; E. cardamomum; southwestern India, Sri Lanka), Renealmia (c 85; tropical Africa, southern Mexico, Central America, the West Indies, tropical South America), Amomum (c 65; northeastern India, southern China, Southeast Asia, West Malesia, New Guinea, northern Queensland), ‘Alpinia’ pro parte, Geostachys (c 25; Southeast Asia, Sumatra, Borneo), Lanxangia (8; Yunnan, northern Thailand, northern Laos, northern Vietnam), Sundamomum (14; peninsular Thailand, the Malay Peninsula, Sumatra, West Java, Borneo), Conamomum (c 10; Indochina, the Malay Peninsula, Sumatra, Borneo), Sulettaria (15; Southeast Asia, Sumatra, Borneo), Geocharis (6; G. aurantiaca, G. fusiformis, G. macrostemon, G. radicalis, G. rubra, G. secundiflora; the Malay Peninsula, Sumatra, Borneo, the Philippines), Epiamomum (6; E. angustipetalum, E. borneense, E. epiphyticum, E. hansenii, E. pungens, E. roseisquamosum; Borneo), Wurfbainia (27; Indochina, the Malay Peninsula to Sulawesi and the Philippines), Meistera (42; India, Sri Lanka, Southeast Asia, Malesia to New Guinea and eastern Queensland), Paramomum (1; P. petaloideum; southwestern China), Plagiostachys (20–25; southern China, Southeast Asia, Nicobar Islands, West Malesia to the Philippines), ’Etlingera’ (>100; tropical Asia from India to southern China, Southeast Asia, Malesia to New Guinea, Queensland and islands in western Pacific; non-monophyletic), Hornstedtia (30–35; tropical Asia from the Himalayas to southern China, Southeast Asia, Malesia to New Guinea, Queensland and Melanesia), Vanoverberghia (1; V. sepulchrei; the Philippines). – UnplacedAlpinioideae Aulotandra (6; A. angustifolia, A. humbertii, A. madagascariensis, A. trialata, A. trigonocarpa: Madagascar; A. kamerunensis: Cameroon), Cyphostigma (1; C. pulchellum; Sri Lanka), Leptosolena (1; L. haenkei; Luzon in the Philippines). – Tropical and southern Africa, Madagascar, the Mascarene Islands, the Seychelles, tropical Asia, tropical Australia and islands in the Pacific, tropical America. Rhizome fleshy. Distichy of leaves perpendicular to rhizome. Pseudolamina in Riedelieae with nectar pits (extrafloral nectaries) on abaxial mid-vein. Lateral staminodia in outer whorl very small or absent, never petaloid. Labellum not connate to filament. Anther crest petaloid or absent. Ovary uni- or trilocular. Styloids present in Aframomum. Flexistyly (style changing direction during anthesis) sometimes present in Alpinieae. Placentation axile, parietal or free central. Fruit in Riedelieae silique-like dehiscing with longitudinal slits, in Alpinieae fleshy or indehiscent. n = 11, 12. – Siliquamomum is sister to Riedelieae (Kress & al. 2002).
Zingiberoideae (Blume) Hassk., Cat. Hort. Bot. Bogor.: 49. Oct 1844 [‘Zingibera’]
33/610–630. Globbeae Meisn., Plant. Vasc. Gen.: Tab. Diagn. 388, Comm. 290. 17-20 Aug 1842. Globba (c 100; India, Sri Lanka, China, Southeast Asia, Malesia to New Guinea and Queensland). – ‘Gagnepainia clade’ Gagnepainia (3; G. godefroyi, G. harmandii, G. thoreliana; Southeast Asia), Hemiorchis (3; H. burmanica, H. pantlingii, H. rhodorrhachis; the Himalayas to Burma). – ‘Caulokaempferiaclade’ Caulokaempferia (c 30; the Himalayas to southern China and Southeast Asia). – Zingibereae Meisn., Plant. Vasc. Gen.: Tab. Diagn. 388, Comm. 290. 17–20 Aug 1842. Camptandra (4; C. gracillima, C. latifolia, C. ovata, C. parvula; West Malesia), Smithatris (2; S. myanmarensis, S. supraneanae; Burma, Thailand; in Curcuma?), Hitchenia (1; H. glauca; Burma; in Curcuma?), Stahlianthus (7; S. andersonii, S. campanulatus, S. involucratus, S. macrochlamys, S. pedicellatus, S. philippianus, S. thorelii; northern India, Burma, southern and southwestern China, Southeast Asia; in Curcuma?), Paracautleya (1; P. bhatii; southern India), Curcuma (c 100; tropical Asia from India and Sri Lanka to southern China and Southeast Asia, Malesia to New Guinea and tropical Australia; paraphyletic?),‘Hedychium (70–80; Madagascar, the Himalayas, southern China, Southeast Asia, West Malesia to the Philippines), Larsenianthus (4; L. arunachalensis, L. assamensis, L. careyanus, L. wardianus; northern India and eastern Himalayas to Burma), Nanochilus (1; N. palembanicum; Sumatra), Roscoea (22; the Himalayas, Burma, western China), Cautleya (2; C. gracilis, C. spicata; the Himalayas), Pommereschea (2; P. lackneri, P. spectabilis; southern China, Burma, Thailand), Rhynchanthus (4; R. beesianus, R. bluthianus, R. johnianus, R. longiflorus; Assam, Yunnan, Burma), Haniffia (3; H. albiflora, H. cyanescens, H. flavescens; Peninsular Thailand, the Malay Peninsula), Curcumorpha (1; C. longiflora; eastern Himalayas; in Boesenbergia?), ’Boesenbergia’ (60–70; India to southern China and Southeast Asia, West Malesia to Borneo; non-monophyletic), Kaempferia (c 35; India to southern China and Southeast Asia, West Malesia to the Philippines, Taiwan), Zingiber (c 100?; India, China, Southeast Asia, Malesia to New Guinea and tropical Australia), Cornukaempferia (3; C. aurantiflora, C. larsenii, C. longipetiolata; Thailand), Distichochlamys (4; D. benenica, D. citrea, D. oriowii, D. rubrostriata; Vietnam), Scaphochlamys (c 30; southern Thailand, the Malay Peninsula, Borneo), Borneocola (8; Borneo), Myxochlamys (2; M. mullerensis, M. nobilis; Borneo), Haplochorema (6; H. decus-sylvae, H. extensum, H. latilabrum, H. magnum, H. pauciflorum: Borneo; H. sumatranum: Sumatra). – Unplaced Zingiberoideae Kedhalia (1; K. flaviflora; Kedah Province on the Malay Peninsula), Newmania (3; N. orthostachys, N. serpens, N. sessilanthera; Vietnam), Laosanthus (1; L. graminifolius; Laos), Parakaempferia (1; P. synantha; Assam), Stadiochilus (1; S. burmanicus; Burma). – Madagascar, the Himalayas, East and tropical Asia, tropical Australia. Rhizome fibrous. Distichy of leaves parallel to rhizome. Lateral staminodia petaloid, adnate to or free from labellum, sometimes connate to filament. Labellum in Zingibereae not connate to filament, in ‘Globbeae’ adnate to filament and forming a tube. Anther crest absent, petaloid, or well developed and wrapped around style (in Zingibereae), or spurred or absent (in ‘Globbeae’). Tapetum sometimes amoeboid-periplasmodial. Pollen grains in Zingiber sulcate. Ovary unilocular or trilocular (sometimes incompletely so). Style with two vascular bundles. Placentation in Zingibereae axile, basal or free columnar, in ‘Globbeae’ parietal. Fruit in Zingibereae fleshy and dehiscent, in ‘Globbeae’ globose and dehiscent. Seeds in Globba and Zingiber carunculate. Exotestal epidermis in ‘Globbeae’ multilayered. Endotesta parenchymatous. Starch grains of perisperm usually compound (in ‘Globbeae’ simple). Embryo in Hedychium long. Steroidal saponins present in Hedychium. n = (8–)10–12(l–14) and higher. – The clade [Globba+Mantisia] forms a sister-group to the remaining Zingiberoideae, and the clade [Gagnepainia+Hemiorchis] is successive sister to [Caulokaempferia+Zingibereae] (Kress & al. 2002).Pommereschea has a parenchymatic endotesta and a long style.
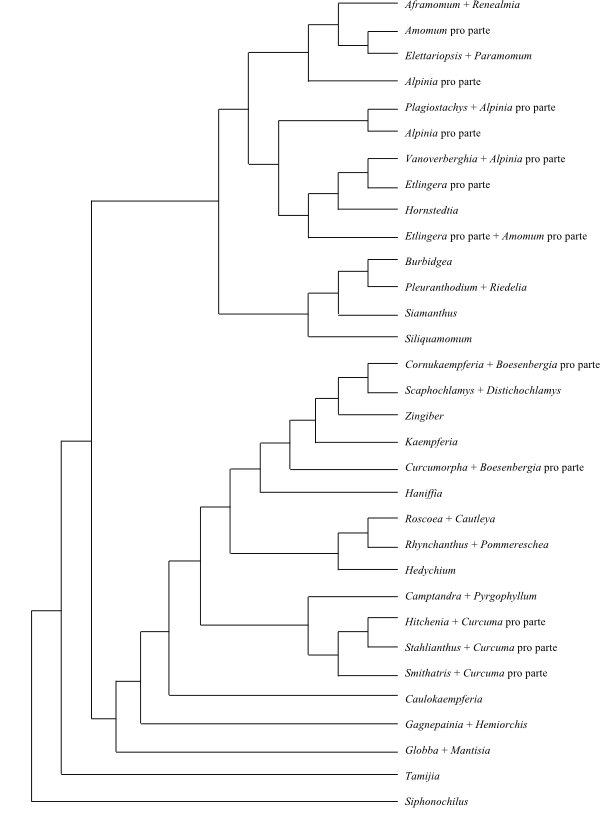 |
Cladogram (one of 980 most-parsimonious) of Zingiberaceae based on DNA sequence data (Kress & al. 2002). |
Literature
Agharkar SP, Bhaduri PN. 1935. Variation in chromosome numbers in Musaceae. – Curr. Sci. 3: 615-617.
Albuquerque E, Braga J, Vieira R. 2013. Morphological characterisation of silica phytoliths in Neotropical Marantaceae leaves. – Plant Syst. Evol. 299: 1659-1670.
Andersson L. 1976. The synflorescence of the Marantaceae. Organization and descriptive terminology. – Bot. Not. 129: 39-48.
Andersson L. 1977. The genus Ischnosiphon (Marantaceae). – Opera Bot. 43: 1-114.
Andersson L. 1981a. Revision of the Thalia geniculata complex (Marantaceae). – Nord. J. Bot. 1: 48-56.
Andersson L. 1981b. The neotropical genera of Marantaceae. Circumscription and relationships. – Nord. J. Bot. 1: 218-245.
Andersson L. 1981c. Revision of Heliconia sect. Heliconia (Musaceae). – Nord. J. Bot. 1: 759-784.
Andersson L. 1984a. Sanblasia, a new genus of the Marantaceae. – Nord. J. Bot. 4: 21-23.
Andersson L. 1984b. Notes on Ischnosiphon (Marantaceae). – Nord. J. Bot. 4: 25-32.
Andersson L. 1984c. The chromosome number of Heliconia (Musaceae). – Nord. J. Bot. 4: 191-193.
Andersson L. 1985a. Revision of Heliconia subgen. Stenochlamys. – Opera Bot. 82: 1-123.
Andersson L. 1985b. Musaceae. – In: Harling G, Sparre B (eds), Flora of Ecuador 22, Swedish Natural Science Research Council, Stockholm, pp. 1-86.
Andersson L. 1985c. The identity of Ctenophrynium K. Schum. (Marantaceae). – Nord. J. Bot. 5: 61-63.
Andersson L. 1986. Revision of Maranta subgen. Maranta (Marantaceae). – Nord. J. Bot. 6: 729-756.
Andersson L. 1992. Revision of Heliconia subgen. Taeniostrobus and subgen. Heliconia (Musaceae-Heliconioideae). – Opera Bot. 111: 1-98.
Andersson L. 1998a. Heliconiaceae. – In: Kubitzki K (ed), The families and genera of vascular plants IV. Flowering plants. Monocotyledons. Alismatanae and Commelinanae (except Gramineae), Springer, Berlin, Heidelberg, New York, pp. 226-230.
Andersson L. 1998b. Marantaceae. – In: Kubitzki K (ed), The families and genera of vascular plants IV. Flowering plants. Monocotyledons. Alismatanae and Commelinanae (except Gramineae), Springer, Berlin, Heidelberg, New York, pp. 278-293.
Andersson L. 1998c. Musaceae. – In: Kubitzki K (ed), The families and genera of vascular plants IV. Flowering plants. Monocotyledons. Alismatanae and Commelinanae (except Gramineae), Springer, Berlin, Heidelberg, New York, pp. 296-301.
Andersson L. 1998d. Strelitziaceae. – In: Kubitzki K (ed), The families and genera of vascular plants IV. Flowering plants. Monocotyledons. Alismatanae and Commelinanae (except Gramineae), Springer, Berlin, Heidelberg, New York, pp. 451-455.
Andersson L, Chase MW. 2001. Phylogeny and classification of Marantaceae. – Bot. J. Linn. Soc. 135: 275-287.
Andersson L, Kennedy H. 1986. Four new species of Calathea (Marantaceae) from French Guiana. – Nord. J. Bot. 6: 447-455.
d’Angremond A. 1915. Parthenokarpie und Samenbildung bei Bananen. – Flora 107: 57-110.
Argent GGG. 1976. The wild bananas of Papua New Guinea. – Notes Roy. Bot. Gard. Edinb. 35: 77-114.
Aristeguieta L. 1961a. Novedades del género Heliconia en Venezuela. – Bol. Soc. Venezolana Ci. Nat. 22: 17-24.
Aristeguieta L. 1961b. El género Heliconia en Venezuela. – Caracas.
Auvray G, Harris DJ, Richardson JE, Newman MF, Särkinen TE. 2010. Phylogeny and dating of Aframomum (Zingiberaceae). – In: Seberg O, Petersen G, Barfod AS, Davis JI (eds), Diversity, phylogeny, and evolution in the monocotyledons, Aarhus University Press, Århus, pp. 287-305.
Baker JG. 1893. A synopsis of the genera and species of Museae. – Ann. Bot. 7: 189-222.
Barreiros H de Souza. 1972a. Heliconia nova brasiliana et varietas. Morfologia e ecologia – dispersão e polinização (Heliconiaceae (Endl.) Nakai). – Rev. Bras. Biol. 32: 205-208.
Barreiros H de Souza. 1972b. Espécies criticas de Heliconia L. I. Ecologia e morfologia (Heliconiaceae). – Rev. Bras. Biol. 32: 297-302.
Barrett CF, Specht CD, Leebens-Mack J,
Stevenson DW, Zomlefer WB, Davis JI. 2014. Resolving ancient radiations: can
complete plastid gene sets elucidate deep relationships among the tropical
gingers (Zingiberales)? – Ann. Bot. 113: 119-133.
Bartlett ME, Specht CD. 2010. Evidence for the involvement of GLOBOSA-like gene duplications and expression divergence in the evolution of floral morphology in the Zingiberales. – New Phytologist 187: 521-541.
Bartlett M, Specht C. 2011. Changes in the expression pattern of the TEOSINTE BRANCHED1-like genes in the Zingiberales provide a mechanism for evolutionary shifts in symmetry across the order. – Amer. J. Bot. 98: 227-243.
Behnke H-D. 1994. Sieve-element plastids, nuclear crystals, and phloem proteins in the Zingiberales. – Bot. Acta 107: 3-11.
Beltran IC, Kam YK. 1984. Cytotaxonomic studies in the Zingiberaceae. – Notes Roy. Bot. Gard. Edinb. 41: 541-559.
Benedict JC, Smith SY, Collinson ME, Leong-Škorničkova J, Specht CD, Fife JL, Marone F, Xiao X, Parkinson DY. 2015. Evolutionary significance of seed structure in Alpinioidee (Zingiberaceae). – Bot. J. Linn. Soc. 178: 441-466.
Benedict JC, Smith SY, Collinson ME, Leong-Škorničkova J, Specht CD, Marone F, Xiao X, Parkinson DY. 2015. Seed morphology and anatomy and its utility in recognizing subfamilies and tribes of Zingiberaceae. – Amer. J. Bot. 102: 1814-1841.
Berry EW. 1925. A banana in the Tertiary of Colombia. – Amer. J. Sci. 10: 530-537.
Berry F, Kress J. 1991. Heliconia. An identification guide. – Smithsonian Institution, Washington, D.C.
Bisson S, Guillemet S, Hamel J-L. 1968. Contribution à l’étude caryo-taxinomique des Scitaminées. – Mém. Mus. Natl. Hist. Nat. Paris, sér. II, sect. B, 18: 59-145.
Boer H de, Newman M, Poulsen AD, Droop AJ, Fér T, Hiên LTT, Hlavatá K, Lamxay V, Richardson JE, Steffen K, Leont-Škorničková J. 2018. Convergent morphology in Alpinieae (Zingiberaceae): recircumscribing Amomum as a monophyletic genus. – Taxon 67: 6-36.
Borchsenius F, Øllgaard B, Suksathan P. 2005. Proposal to conserve the name Stachyphrynium against Phyllodes (Marantaceae). – Taxon 54: 833-834.
Borchsenius F, Suárez LSS, Prince LM. 2012. Molecular phylogeny and redefined generic limits of Calathea (Marantaceae). – Syst. Bot. 37: 620-635.
Bouharmont J. 1963. Évolution de l’ovule fécondé chez Musa acuminata Colla subsp. burmannica Simmonds. – Cellule 63: 262-279.
Box MS, Rudall PJ. 2006. Floral structure and ontogeny in Globba (Zingiberaceae). – Plant Syst. Evol. 258: 107-122.
Braun J, Schumann K. 1889. Zingiberaceae. – Mitt. Forschungsreis. Gelehrten Deutsch. Schutzgeb. 2: 151-153.
Burtt BL. 1972. General introduction to papers on Zingiberaceae. – Notes Roy. Bot. Gard. Edinb. 31: 155-165.
Burtt BL, Smith RM. 1972a. Tentative keys to the subfamilies, tribes, and genera of Zingiberaceae. – Notes Roy. Bot. Gard. Edinb. 31: 171-176.
Burtt BL, Smith RM. 1972b. Key species in the taxonomic history of Zingiberaceae. – Notes Roy. Bot. Gard. Edinb. 31: 177-227.
Burtt BL, Smith RM. 1986. Etlingera: the inclusive name for Achasma, Geanthus and Nicolaia (Zingiberaceae). – Notes Roy. Bot. Gard. Edinb. 43: 235-241.
Carlquist SJ, Schneider EL. 1998. Origins and nature of vessels in monocotyledons 3. Lowiaceae, with comments on rhizome anatomy. – Blumea 43: 219-224.
Carlquist SJ, Schneider EL. 2010. Origins and nature of vessels in monocotyledons 11. Primary xylem microstructure, with examples from Zingiberales. – Intern. J. Plant Sci. 171: 258-266.
Chakravorti AK. 1960. Idiogram studies with special reference to chromosome-nucleolus relationship and its bearing on the cytogenetics of Heliconia. – Nucleus 3: 225-250.
Cheesman EE. 1947. Classification of the bananas. – Kew Bull. 2: 97-117.
Cheesman EE, Larter LNH. 1935. Genetical and cytological studies of Musa III. Chromosome numbers in the Musaceae. – J. Genet. 30: 31-52.
Chen ST, Smith SY. 2013. Phytolith variability in Zingiberales: a tool for the reconstruction of past tropical vegetation. – Palaeogeogr. Palaeoclim. Palaeoecol. 370: 1-12.
Chen Z-Y. 1989. Evolutionary patterns in cytology and pollen structure of Asian Zingiberaceae. – In: Holm-Nielsen LB, Nielsen IC, Balslev H (eds), Tropical forests. Botanical dynamics, speciation and diversity, Academic Press, London, pp. 185-199.
Chevalier A. 1917. Zingiberaceae. – Bull. Soc. Bot. France 61, sér. 4, 14, Mém. 8: 304-305.
Chevalier A. 1920. Exploration botanique de l’Afrique occidentale française 1: Zingiberaceae. – Lechevallier, Paris, pp. 627-628.
Christelova P, Valarik M, Hribova E, Lange E de, Dolezel J. 2011. A multi gene sequence-based phylogeny of the Musaceae (banana) family. – BMC Ecol. Biol. 11: 103.
Ciciarelli M de las Mercedes. 1995. Canna variegatifolia Ciciarelli sp. nov. (Cannaceae: Zingiberales), una especie nueva para Argentina. – Rev. Mus. La Plata, Secc. Bot. 14: 333-341.
Ciciarelli M de las Mercedes. 2007. Canna ascendens (Cannaceae), una nueva especie de la provincia de Buenos Aires y comentarios sobre otras especies Argentinas de este género. – Darwiniana 45: 188-200.
Claßen-Bockhoff R. 1991. Untersuchungen zur Konstruktion des Bestäubungsapparates von Thalia geniculata (Marantaceae). – Bot. Acta 194: 183-193.
Claßen-Bockhoff R, Heller A. 2008. Floral synorganization and secondary pollen presentation in four Marantaceae from Costa Rica. – Intern. J. Plant Sci. 169: 745-760.
Clausager K, Borchsenius F. 2003. The Marantaceae of Sabah, Northern Borneo. – Kew Bull. 58: 647-678.
Cowley EJ. 1982. A revision of Roscoea (Zingiberaceae). – Kew Bull. 36: 747-777.
Cowley J. 1998. Two new species of Boesenbergia (Zingiberaceae) from Brunei Darussalam. – Kew Bull. 53: 623-629.
Cowley J. 1999. Two new species of Plagiostachys (Zingiberaceae) from Borneo. – Kew Bull. 54: 139-146.
Cowley J. 2000. Three new gingers from Borneo. – Kew Bull. 55: 669-678.
Cron GV, Pirone C, Bartlett M, Kress WJ, Specht C. 2012. Phylogenetic relationships and evolution in the Strelitziaceae (Zingiberales). – Syst. Bot. 37: 606-619.
Cronquist A. 1978. The Zingiberidae, a new subclass of Liliopsida (monocotyledons). – Brittonia 30: 505.
Daghlian CP. 1982. Zingiberalean leaves from the lower Eocene of Texas. – Bot. Soc. Amer. Misc. Publ. 162: 57.
Daniels GS, Stiles FG. 1979. The Heliconia taxa of Costa Rica. Keys and descriptions. – Brenesia 15 [Suppl.]: 1-149.
Desler B. 1994. Vergleichende Untersuchung der Anatomie und Morphologie von Scitamineen-Samen. – Diploma Thesis, Universität Kaiserslautern, Germany.
Dhetchuvi M-M. 1996. Taxonomie et phytogeographie des Marantaceae et des Zingiberaceae de l’Afrique Centrale (Gabon, Congo, Zaire, Rwanda et Burundi). – Ph.D. diss., Université Libre de Bruxelles, Belgium.
Eichler AW. 1884. Beiträge zur Morphologie und systematic der Marantaceen. – Abh. K. Preuss. Akad. Wissensch. Berlin 1883: 1-97.
Fahn A. 1953. The origin of the banana inflorescence. – Kew Bull. 1953: 299-306.
Fahn A, Benouaiche P. 1979. Ultrastructure, development, and secretion in the nectary of banana flowers. – Ann. Bot., N. S., 44: 85-93.
Fahn A, Kotler M. 1972. The development of the flower and nectary of the dwarf Cavedish banana. – Adv. Plant Morph. 1972: 153-169.
Fauré S, Noyer J-L, Carreel F, Horry J-P, Bakry F, Lanaud C. 1994. Maternal inheritance of chloroplast genome and paternal inheritance of mitochondrial genome in bananas (Musa acuminata). – Curr. Genet. 25: 265-269.
Fisher JB. 1976. Development of dichotomous branching and axillary buds in Strelitzia (Monocotyledonae). – Can. J. Bot. 54: 578-592.
Fisher JB. 1978. Leaf-opposed buds in Musa: their development and a comparison with allied monocotyledons. – Amer. J. Bot. 65: 784-791.
Freeman B, Turner DW. 1985. The epicuticular waxes on the organs of different varieties of banana (Musa spp.) differ in form, chemistry and concentration. – Aust. J. Bot. 33: 393-408.
Friedrich WL, Koch BE. 1970. Comparison of fruits and seeds of fossil Spirematospermum (Zingiberaceae) with those of living Cenolophon. – Bull. Geol. Soc. Denmark 20: 192-195.
Friedrich WL, Koch BE. 1972. Der Arillus der tertiären Zingiberaceae Spirematospermum wetzleri. – Lethaia 5: 47-60.
Friedrich WL, Strauch F. 1975 [1976]. Der Arillus der Gattung Musa. – Bot. Not. 128: 339-349.
Friis EM. 1988. Spirematospermum chandlerae sp. nov., an extinct Zingiberacea from the North American Cretaceous. – Tertiary Res. 9: 7-12.
Frost SK, Frost PGH. 1981. Sunbird pollination of Strelitzia nicolai. – Oecologia 49: 379-384.
Funakoshi H, Ohashi H. 2000. Vanoverberghia sasakiana H. Funk & H. Ohashi (Zingiberaceae), a new species and a new generic record for the Flora of Taiwan. – Taiwania 45: 270-275.
Gade D. 1966. Achira, the edible Canna, its cultivation and use in the Peruvian Andes. – Econ. Bot. 20: 407-415.
Gagnepain MF. 1901. Révision des genres Mantisia et Globba (Zingibérées) de l’herbier du muséum. – Bull. Soc. Bot. France, sér. IV, 1:. 201-216.
Gagnepain MF. 1902a. Zingibéracées nouvelles de l’Herbier du Muséum. – Bull. Soc. Bot. France, sér. IV, 49: 93-97.
Gagnepain MF. 1902b. Les Zingibéracées du continent africain dans l’herbier du Muséum. – Bull. Soc. Hist. Nat. 15: 16-25.
Gagnepain MF. 1903a. Zingibéracées nouvelles de l’herbier du muséum. – Bull. Soc. Bot. France, sér. IV, 50: 160-165.
Gagnepain MF. 1903b. Zingibéracées de l’herbier du muséum. – Bull. Soc. Bot. France, sér. IV, 50: 257-263.
Gagnepain MF. 1904. Zingibéracées et Marantacées nouvelles de l’herbier du muséum. – Bull. Soc. Bot. France 51: 164-182.
Gagnepain MF. 1907. Zingibéracées nouvelles des l’herbier du muséum. – Bull. Soc. Bot. France, sér. IV, 7: 111-115.
Gao J-Y, Zhang L, Deng X-B, Ren P-Y, Kong J-J, Li Q-J. 2004. The floral biology of Curcumorpha longiflora (Zingiberaceae): a ginger with two-day flowers. – Amer. J. Bot. 91: 289-293.
Gatin CL. 1908. Recherches anatomiques sur l’embryon et la germination des Cannacées et des Musacées. – Ann. Sci. Nat. Bot. 8: 113-146.
Gawel NJ, Jarret RL. 1991. Chloroplast DNA restriction fragment length polymorphisms (RFLPs) in Musa species. – Theor. Appl. Genet. 81: 783-786.
Gawel NJ, Jarret RL, Whittemore AP. 1992. Restriction fragment length polymorphism (RFLP)-based phylogenetic analysis of Musa. – Theor. Appl. Genet. 84: 286-290.
Geri C, Ng FSP. 2005. Musa lokok (Musaceae), a new species of banana from Bario, Borneo. – Gard. Bull. Singapore 57: 279-283.
Goth K. 1986. Erster Nachweis von Spirematospermum-Samen aus der Oberkreide von Kössen in Tirol. – Courier Forschungsinst. Senckenberg 86: 171-175.
Govaerts R. 2013. Hellenia Retz., the correct name for Cheilocostus C. D. Specht (Costaceae). – Phytotaxa 151: 63-64.
Graven P, de Koster CG, Boon JJ, Bouman F. 1996. Structure and molecular composition of the seed coat of Musaceae. – Ann. Bot. 77: 105-122.
Griggs RF. 1903. On some species of Heliconia. – Bull. Torrey Bot. Club 30: 641-664.
Griggs RF. 1904. Two new species of American wild bananas with a revision of the generic name. – Bull. Torrey Bot. Club 31: 445-447.
Griggs RF. 1915. Some new species and varieties of Bihai. – Bull. Torrey Bot. Club 42: 315-330.
Grootjen CJ. 1983. Development of ovule and seed in Marantaceae. – Acta Bot. Neerl. 32: 69-86.
Grootjen CJ, Bouman F. 1981. Development of the ovule and seed in Costus cuspidatus (N. E. Br.) Maas (Zingiberaceae), with special reference to the formation of the operculum. – Bot. J. Linn. Soc. 83: 27-39.
Grootjen CJ, Bouman F. 1988. Seed structure in Cannaceae: taxonomic and ecological implications. – Ann. Bot., N. S., 61: 363-371.
Hagberg M, Eriksson R. 2011. New names in Monotagma. – Phytotaxa 20: 1-25.
Häkkinen M. 2005. Musa azizii, a new Musa species (Musaceae) from northern Borneo. – Acta Phytotaxon. Geobot. 56: 27-31.
Häkkinen M. 2006a. Musa lawitiensis Nasution & Supard. (Musaceae) and its intraspecific taxa in Borneo. – Adansonia, sér. III, 28: 55-65.
Häkkinen M. 2006b. Musa barioensis, a new Musa species (Musaceae) from northern Borneo. – Acta Phytotaxon. Geobot. 57: 55-60.
Häkkinen M, Meekiong K. 2004. A new species of the wild banana genus, Musa (Musaceae), from Borneo. – Syst. Biodivers. 2: 169-173.
Häkkinen M, Väre H. 2008. Typification and check-list of Musa L. names (Musaceae) with nomenclatural notes. – Adansonia, sér. III, 30: 63-112.
Häkkinen M, Wang H. 2008. Musa zaifui sp. nov. (Musaceae) from Yunnan, China. – Nord. J. Bot. 26: 42-46.
Hallé N. 1979. Architecture du rhizome chez quelques Zingibéracées d'Afrique et d’Océanie. – Adansonia, sér. 2, 19, 2: 127-144.
Harris DJ, Poulsen AD, Frimodt-Møller C, Preston J, Cronk QCB. 2000. Rapid radiations in Aframomum (Zingiberaceae): evidence from nuclear ribosomal DNA internal transcribed spacer (ITS) sequences. – Edinburgh J. Bot. 57: 377-395.
Harris DJ, Newman MF, Hollingsworth ML, Möller M, Clark A. 2006. The phylogenetic position of Aulotandra Gagnep. (Zingiberaceae). – Nord. J. Bot. 23: 725-734.
Hema PS, Nair MS. 2009. Flavonoids and other constituents from the rhizomes of Alpinia calcarata. – Biochem. Syst. Ecol. 37: 52-54.
Hermann M. 1996. Starch noodles from edible canna. – In: Janick J (ed), Progress in new crops, ASHS Press, Arlington.
Hesse M, Waha M. 1983. The fine structure of the pollen wall in Strelitzia reginae (Musaceae). – Plant Syst. Evol. 141: 285-298.
Hickey LH, Peterson RK. 1978. Zingiberopsis, a fossil genus of the ginger family from Late Cretaceous to Early Eocene sediments of Western Interior North America. – Can. J. Bot. 56: 1136-1152.
Hodge WH. 1941. The type species of Heliconia. – Contr. Gray Herb. Harvard Univ. 135: 134-137.
Hölscher D, Schneider B. 1998. Phenylphenalenones from Ensete ventricosum. – Phytochemistry 49: 2155-2157.
Hölscher D, Schneider B. 2000. Phenalenones from Strelitzia reginae. – J Nat. Prod. 63: 1027-1028.
Holttum RE. 1950. The Zingiberaceae of the Malay Peninsula. – Gard. Bull. Straits Settlem. (Singapore) 13: 1-249.
Holttum RE. 1951. The Marantaceae of Malaya. – Gard. Bull. Straits Settlem. (Singapore) 13: 254-296.
Holttum RE. 1970. The genus Orchidantha (Lowiaceae). – Gard. Bull. (Singapore) 25: 239-246.
Holttum RE. 1974. A commentary on comparative morphology in Zingiberaceae. – Gard. Bull. (Singapore) 27: 155-165.
Honing JA. 1939. Canna crosses IV. – Genetica 15: 23-47.
Horvitz CC, Beattie AJ. 1980. Ant dispersal of Calathea (Marantaceae) seeds by carnivorous Ponerines (Formicidae) in a tropical rain forest. – Amer. J. Bot. 67: 321-326.
Humphrey JESD. 1896. The development of the seed in the Scitamineae. – Ann. Bot. 10: 1-40.
Hussin KH, Seng CT, Ibrahim H, Gen WQ, Ping LJ, Nian L. 2000. Comparative leaf anatomy of Alpinia Roxb. species (Zingiberaceae) from China. – Bot. J. Linn. Soc. 133: 161-180.
Ibrahim H, Larsen K. 1995. A new species of Globba (Zingiberaceae) from peninsular Malaysia. – Nord. J. Bot. 15: 157-159.
Iles WJD, Sass C, Lagomarsino L, Benson-Martin G, Driscoll H, Specht CD. 2017. The phylogeny of Heliconia (Heliconiaceae) and the evolution of floral presentation. – Mol. Phylogen. Evol. 117: 150-167.
Isobe M, Hashimoto K. 1994. The chromosome count of nine taxa in Musa and its allied genus Musella. – Bull. Hiroshima Bot. Gard. 15: 7-11.
Jain RK. 1964. Studies of Musaceae I. Musa cardiosperma sp. nov., a fossil banana fruit from the Deccan Intertrappean series, India. – The Palaeobotanist 12: 45-58.
Jain RK. 1965. Studies in Musaceae III. Fossil records of Musaceae and the origin of bananas. – Proc. Indian Acad. Sci. 6: 170-179.
Jain SK, Prakash V. 1995. Zingiberaceae in India: phytogeography and endemism. – Rheedea 5: 166.
Jarret RL (ed). 1990. Identification of genetic diversity in the genus Musa. – INIBAP, Montpellier.
Jarret RL, Gawel N, Whittemore A, Sharrock S. 1992. RFLP-based phylogeny of Musa species in Papua New Guinea. – Theor. Appl. Genet. 84: 579-584.
Jesson LK, Barrett SCH. 2003. The comparative biology of mirror-image flowers. – Intern. J. Plant Sci. 164(Suppl.): S237-S249.
Johansen LB. 2005. Phylogeny of Orchidantha (Lowiaceae) and the Zingiberales based on six DNA regions. – Syst. Bot. 30: 106-117.
Johnson CT. 1977. ‘N anatomiese ondersoek van die blom van Strelitzia reginae Banks. – J. South Afr. Bot. 43: 81-91.
Jongkind CCH. 2008. Two new species of Hypselodelphys (Marantaceae) from West Africa. – Adansonia, sér. III, 30: 57-62.
Jonker-Verhoef AME, Jonker FP. 1955. Notes on the Marantaceae of Suriname. – Acta Bot. Neerl. 4: 172-182.
Kaewsri W, Paisooksantivatana Y. 2007. Morphology and palynology of Amomum Roxb. in Thailand. – Gard. Bull. Singapore 59: 105-112.
Kamelina OP. 1990. The development of male and female embryonic structures in the Marantaceae family. – Bot. Žurn. 75: 480-483. [In Russian]
Kamo T, Hirai N, Iwami K, Fujioka D, Ohigashi H. 2001. New phenylphenalenones from banana fruit. – Tetrahedron 57: 7649-7656.
Kamo T, Kato N, Hirai N, Tsuda M, Fujioka D, Ohigashi H. 1998. Phenylphenalenone-type phytoalexins from unripe bungulan banana fruit. – Biosci. Biotechn. Biochem. 62: 95-101.
Kay KM, Reeves PA, Olmstead RG, Schemske DW. 2005. Rapid speciation and the evolution of hummingbird pollination in neotropical Costus subgenus Costus (Costaceae): evidence from nrDNA ITS and ETS sequences. – Amer. J. Bot. 92: 1899-1910.
Kennedy H. 1973. Notes on Central American Marantaceae I. New species and records from Panamá and Costa Rica. – Ann. Missouri Bot. Gard. 60: 413-426.
Kennedy H. 1974. Pollination and systematics of the “close-flowered” species of Calathea (Marantaceae). – Ph.D. diss., University of California, Davis, California.
Kennedy H. 1975 [1976]. Notes on Central American Marantaceae II. New species from Panamá and Costa Rica. – Bot. Not. 128: 312-322.
Kennedy H. 1977a. Unusual floral morphology in a high altitude Calathea (Marantaceae). – Brenesia 12/13: 1-9.
Kennedy H. 1977b. New species of Calathea from Panama and Costa Rica. – Bot. Not. 129: 349-357.
Kennedy H. 1977c. An unusual flowering strategy and new species in Calathea. – Bot. Not. 130: 333-339.
Kennedy H. 1978a. Systematics and pollination of the “closed-flowered” species of Calathea (Marantaceae). – Univ. Calif. Publ. Bot. 71: 1-90.
Kennedy H. 1978b. Calathea elegans and C. spiralis – new Panamanian Marantaceae. – Bot. Not. 131: 349-354.
Kennedy H. 1985. New Ecuadorian species in Calathea series Nudiscapae (Maranthaceae). – Can. J. Bot. 63: 1141-1149.
Kennedy H. 1986a. Additional Ecuadorian species in Calathea series Comosae (Marantaceae). – Nord. J. Bot. 6: 143-150.
Kennedy H. 1986b. New useful and colourfully-bracted species of Calathea (Marantaceae) from Ecuador. – Nord. J. Bot. 6: 457-461.
Kennedy H. 1986c. Notes on Malaysian Marantaceae and the identity of Heliconia triumphans. – Kew Bull. 41: 725-731.
Kennedy H. 2000. Diversification in pollination mechanisms in the Marantaceae. – In: Wilson KL, Morrison AA (eds), Monocots: systematics and evolution, CSIRO, Melbourne, pp. 335-343.
Kennedy H, Andersson L, Hagberg M. 1988. 224. Marantaceae. – In: Harling G, Andersson L (eds), Flora of Ecuador 32, Nord. J. Bot., Copenhagen, pp. 13-191.
Khoshoo TN. 1972. Evolution of garden cannas. – In: Vida G (ed), Evolution in plants, Symp. Biol. Hungar., Vol. 12, pp. 93-101.
Kiew KY. 1982. The genus Elettariopsis (Zingiberaceae) in Malaya. – Notes Roy Bot. Gard. Edinb. 40: 139-152.
Kirchoff BK. 1983a. Floral organogenesis in five genera of the Marantaceae and in Canna (Cannaceae). – Amer. J. Bot. 70: 508-523.
Kirchoff BK. 1983b. Allometric growth of the flowers in the five genera of the Marantaceae and in Canna (Cannaceae). – Bot. Gaz. (Crawfordsville) 144: 110-118.
Kirchoff BK. 1986. Inflorescence structure and development in the Zingiberales: Thalia geniculata (Marantaceae). – Can. J. Bot. 64: 859-864.
Kirchoff BK. 1988a. Inflorescence and flower development in Costus scaber (Costaceae). – Can. J. Bot. 66: 339-345.
Kirchoff BK. 1988b. Floral ontogeny and evolution in the ginger group of the Zingiberales. – In: Leins P, Tucker SC, Endress PK (eds), Aspects of floral development, J. Cramer, Berlin, pp. 45-56.
Kirchoff BK. 1991. Homeosis in the flowers of the Zingiberales. – Amer. J. Bot. 78: 833-837.
Kirchoff BK. 1992. Ovary structure and anatomy in the Heliconiaceae and Musaceae (Zingiberales). – Can. J. Bot. 70: 2490-2508.
Kirchoff BK, Kennedy H. 1985. Foliar, non-structural nectaries in the Marantaceae. – Can. J. Bot. 63: 1785-1788.
Kirchoff BK, Kunze H. 1995. Inflorescence and floral development in Orchidantha maxillarioides (Lowiaceae). – Intern. J. Plant Sci. 156: 159-171.
Kirchoff BK, Lagomarsino LP, Newman WH, Bartlett ME, Specht CD. 2009. Early floral development of Heliconia latispatha (Heliconiaceae), a key taxon for understanding the evolution of flower development in the Zingiberales. – Amer. J. Bot. 96: 580-593.
Koch BE, Friedrich WL. 1971. Früchte und Samen von Spirematospermum aus der miozänen Fasterholt-Flora in Dänemark. – Palaeontographica, Ser. B, 136: 1-46.
Koechlin J. 1964. Scitaminales: Musacées, Strélitziacées, Zingibéracées, Cannacées, Marantacées. – In: Aubréville A (ed), Flore du Gabon 9, Muséum National d’Histoire Naturelle, Paris, pp. 21-32.
Koechlin J. 1965. Scitaminales: Musacées, Strélitziacées, Zingibéracées, Cannacées, Marantacées. – In: Aubréville A (ed), Flore du Cameroun 4, Muséum National d’Histoire Naturelle, Paris, pp. 27-41.
Kress WJ. 1983a. Self-incompatibility in Central American Heliconia. – Evolution 37: 735-744.
Kress WJ. 1983b. Crossability barriers in neotropical Heliconia. – Ann. Bot., N. S., 52: 131-147.
Kress WJ. 1984. Systematics of Central American Heliconia (Heliconiaceae) with pendant inflorescences. – J. Arnold Arbor. 65: 429-532.
Kress WJ. 1985a. Bat pollination of an Old World Heliconia. – Biotropica 17: 302-308.
Kress WJ. 1985b. Pollination and reproduction biology of Heliconia. – In: D’Arcy WG, Correa A (eds), The botany and natural history of Panama, Monogr. Syst. Bot. Missouri Bot. Gard. 10: 267-271.
Kress WJ. 1986. Exineless pollen structure and pollination systems of tropical Heliconia (Heliconiaceae). – In: Blackmore S, Ferguson IK (eds), Pollen and spores. Form and function, Academic Press, London, pp. 329-345.
Kress WJ. 1990a. The phylogeny and classification of the Zingiberales. – Ann. Missouri Bot. Gard. 77: 698-721.
Kress WJ. 1990b. The taxonomy of Old World Heliconia (Heliconiaceae). – Allertonia 6: 1-58.
Kress WJ. 1993. Morphology and floral biology of Phenakospermum (Strelitziaceae), an arborescent herb of the Neotropics. – Biotropica 25: 290-300.
Kress WJ. 1995. Phylogeny of the Zingiberanae: morphology and molecules. – In: Rudall PJ, Cribb PJ, Cutler DF, Humphries CJ (eds), Monocotyledons: systematics and evolution, Royal Botanic Gardens, Kew, pp. 443-460.
Kress WJ, Larsen K. 2001. Smithatris, a new genus of Zingiberaceae from Southeast Asia. – Syst. Bot. 26: 226-230.
Kress WJ, Specht CD. 2005. Between Cancer and Capricorn: phylogeny, evolution and ecology of the primarily tropical Zingiberales. – Biol. Skr. 55: 459-478. [In: Friis I, Balslev H (eds), Proceedings of a symposium on plant diversity and complexity patterns. Local, regional and global dimensions, Danish Academy of Sciences and Letters, Copenhagen, pp. 459-478]
Kress WJ, Specht CD. 2006. The evolutionary and biogeographic origin and diversification on the tropical monocot order Zingiberales. – In: Columbus JT, Friar EA, Porter JM, Prince LM, Simpson MG (eds), Monocots: comparative biology and evolution. Excluding Poales, Rancho Santa Ana Botanical Garden, Claremont, California. – Aliso 22: 621-632.
Kress WJ, Stone DE. 1982. Nature of the sporoderm in monocotyledons, with special reference to the pollen grains of Canna and Heliconia. – Grana 21: 129-148.
Kress WJ, Stone DE. 1983. Morphology and phylogenetic significance of exine-less pollen of Heliconia (Heliconiaceae). – Syst. Bot. 8: 149-167.
Kress WJ, Stone DE, Sellers SC. 1978. Ultrastructure of exineless pollen: Heliconia (Heliconiaceae). – Amer. J. Bot. 65: 1064-1076.
Kress WJ, Schatz GE, Andrianifahanana M, Morland HS. 1994. Pollination of Ravenala madagascariensis (Strelitziaceae) by lemurs in Madagascar: evidence for an archaic coevolutionary system? – Amer. J. Bot. 81: 542-551.
Kress WJ, Prince LM, Hahn WJ, Zimmer EA. 2001. Unraveling the evolutionary radiation of the families of the Zingiberales using morphological and molecular evidence. – Syst. Biol. 50: 926-944.
Kress WJ, Prince LM, Williams KJ. 2002. The phylogeny and a new classification of the gingers (Zingiberaceae): evidence from molecular data. – Amer. J. Bot. 89: 1682-1696.
Kress WJ, Liu A-Z, Newman M, Li Q-J. 2005. The molecular phylogeny of Alpinia (Zingiberaceae): a complex and polyphyletic genus of gingers. – Amer. J. Bot. 92: 167-178.
Kress WJ, Newman M, Poulsen MF, Specht C. 2007. An analysis of generic circumscriptions in tribe Alpinieae (Alpinioideae: Zingiberaceae). – Gard. Bull. (Singapore) 59: 113-128.
Kress WJ, Mood JD, Sabu M, Prince LM, Dey S, Sanoj E. 2010. Larsenianthus, a new Asian genus of gingers (Zingiberaceae) with four species. – PhytoKeys 1: 15-32.
Kronestedt E, Bystedt P-A. 1981. Thread-like formations in the anthers of Strelitzia reginae. – Nord. J. Bot. 1: 523-529.
Kronestedt E, Robards A. 1985. Nectar secretion by the flower of Strelitzia reginae. – Roy. Microscop. Soc. 20: BM 20-21.
Kronestedt E, Walles B. 1983. Wall growth and separation of thread-forming cells in Strelitzia reginae. – J. Ult. Res. 85: 110.
Kronestedt E, Walles B. 1984. Secretory cells of the transmitting tissue in the pistil of Strelitzia reginae. – J. Ult. Res. 88: 303-304.
Kronestedt E, Walles B. 1986. Anatomy of the Strelitzia reginae flower (Strelitziaceae). – Nord. J. Bot. 6: 307-320.
Kubitzki K. 1998. Cannaceae. – In: Kubitzki K (ed), The families and genera of vascular plants IV. Flowering plants. Monocotyledons. Alismatanae and Commelinanae (except Gramineae), Springer, Berlin, Heidelberg, New York, pp. 103-106.
Kunze H. 1984. Vergleichende Studien an Cannaceen- und Marantaceenblüten. – Flora 175: 301-318.
Kunze H. 1985. Die Infloreszenzen der Marantaceen und ihr Zusammenhang mit dem Typus der Zingiberales-Synfloreszenz. – Beitr. Biol. Pflanzen 60: 93-140.
Kunze H. 1986. Infloreszenz- und Blütenmorphologie von Orchidantha maxillarioides (Ridl.) K. Schum. (Lowiaceae). – Beitr. Biol. Pflanzen 61: 221-234.
Lane IE. 1955. Genera and generic relationships in Musaceae. – Mitt. Bot. Staatssamml. München 2(13): 114-131.
Langhe E De, Pillay M, Tenkouano A, Swennen R. 2005. Integrating morphological and molecular taxonomy in Musa: the African plantains (Musa spp. AAB group). – Plant Syst. Evol. 255: 225-236.
Larsen K. 1961. New species of Veratrum and Orchidantha from Thailand and Laos. – Bot. Tidsskr. 56: 345-350.
Larsen K. 1962a. Studies in Zingiberaceae. The genus Geostachys in Thailand. – Bot. Tidsskr. 58: 43-46.
Larsen K. 1962b. Studies in Zingiberaceae III. On a new species of Kaempferia from Thailand and its relatives. – Bot. Tidsskr. 58: 191-203.
Larsen K. 1964. Studies on Zingiberaceae IV: Caulokaempferia, a new genus. – Bot. Tidsskr. 60: 165-179.
Larsen K. 1965. Studies in Zingiberaceae V. Hedychium. Some new Indo-Chinese taxa. – Bot. Tidsskr. 61: 71-79.
Larsen K. 1966. Chromosome cytology and relationship of the Lowiaceae. – Nat. Hist. Bull. Siam. Soc. 21: 21-24.
Larsen K. 1972a. Studies in the genus Globba in Thailand. – Notes Roy. Bot. Gard. Edinb. 31: 229-241.
Larsen K. 1972b. Studies in the genus Globba in Thailand. Appendix. – Notes Roy. Bot. Gard. Edinb. 31: 241.
Larsen K. 1973. A new species of Orchidantha (Lowiaceae) from Vietnam. – Adansonia, sér. II, 13: 481-482.
Larsen K. 1981. Notes on some species of Achasma (Zingiberaceae). – Nord. J. Bot. 1: 45-47.
Larsen K. 1993. A new species of Orchidantha (Lowiaceae) from Borneo. – Nord. J. Bot. 13: 285-288.
Larsen K. 1996. Lowiaceae. – In: Kalkman C et al. (eds), Flora Malesiana I, 12(2), Flora Malesiana Foundation, Rijksberbarium/Hortus Botanicus, Leiden, pp. 763-774.
Larsen K. 1997. Further studies in the genus Boesenbergia (Zingiberaceae). – Nord. J. Bot. 17: 361-366.
Larsen K. 1998a. Costaceae. – In: Kubitzki K (ed), The families and genera of vascular plants IV. Flowering plants. Monocotyledons. Alismatanae and Commelinanae (except Gramineae), Springer, Berlin, Heidelberg, New York, pp. 128-132.
Larsen K. 1998b. Lowiaceae. – In: Kubitzki K (ed), The families and genera of vascular plants IV. Flowering plants. Monocotyledons. Alismatanae and Commelinanae (except Gramineae), Springer, Berlin, Heidelberg, New York, pp. 275-277.
Larsen K, Jenjittikul T. 2001. Laosanthus, a new genus of Zingiberaceae from Laos. – Nord. J. Bot. 21: 135-138.
Larsen K, Mood J. 1998. Siamanthus, a new genus of Zingiberaceae from Thailand. – Nord. J. Bot. 18: 393-397.
Larsen K, Smith RM. 1972. Notes on Caulokaempferia. – Notes Roy. Bot. Gard. Edinb. 31: 287-295.
Larsen K, Lock JM, Maas H, Maas PJM. 1998. Zingiberaceae. – In: Kubitzki K (ed), The families and genera of vascular plants IV. Flowering plants. Monocotyledons. Alismatanae and Commelinanae (except Gramineae), Springer, Berlin, Heidelberg, New York, pp. 474-495.
Leong-Škorničková J, Lý N-S, Poulsen AD, Tosh J, Forrest A. 2011. Newmania: a new ginger genus from central Vietnam. – Taxon 60: 1386-1396.
Lescot M, Piffanelli P, Ciampi AY, Ruiz M, Blanc G, Leebens-Mack J, Silva FR da; Santos CMR, d’Hont A, Garsmeur O, Vilarinhos AD, Kanamori H, Matsumoto T. Ronning CM, Cheung F, Haas BJ, Althoff R, Arbogast T, Hine E, Pappas GJ Jr, Sasaki T, Souza MT Jr, Miller RNG, Glaszmann J-C, Town CD. 2008. Insights into the Musa genome: syntenic relationships to rice and between Musa species. – BMC Genomics 9: 58.
Ley AR, Claßen-Bockhoff R. 2009. Pollination syndromes in African Marantaceae. – Ann. Bot. 104: 41-56.
Ley AR, Claßen-Bockhoff R. 2010. Parallel evolution in plant pollinator interaction in African Marantaceae. – In: Burgh X van der, Maesen J van der, Onana JM (eds), Systematics and conservation of African plants, Royal Botanic Gardens, Kew, pp. 847-854.
Ley AR, Claßen-Bockhoff R. 2011a. Ontogenetic and phylogenetic diversification in Marantaceae. – In: Wanntorp L, Ronse De Craene LP (eds), Flowers on the Tree of Life, Syst. Assoc. Spec. Vol. 80, Cambridge University Press, Cambridge, pp. 236-255.
Ley AR, Claßen-Bockhoff R. 2011b. Evolution in African Marantaceae – evidence from phylogenetic, ecological and morphological studies. – Syst. Bot. 36: 277-290.
Ley AR, Claßen-Bockhoff R. 2012. Five new species of Marantaceae endemic to Gabon. – Adansonia 34: 37-52.
Li H-W. 1978. The Musaceae of Yunnan. – Acta Phytotaxon. Sin. 16: 54-64. [in Chinese]
Liang Y-H. 1988. Pollen morphology of the family Zingiberaceae in China – pollen types and their significance in the taxonomy. – Acta Phytotaxon. Sin. 26: 265-281. [In Chinese with English abstract]
Liao J-P, Wu Q-G. 2000. A preliminary study of the seed anatomy of Zingiberaceae. – Bot. J. Linn. Soc. 134: 287-300.
Liao J-P, Tang Y-J, Ye X-L, Wu Q-G. 2004. Seed anatomy of species in banana families of Zingiberales and its systematic significance. – J. Trop. Subtrop. Bot. 12: 291-297. [In Chinese]
Lim S-N. 1972. Cytogenetics and taxonomy of the genus Globba L. (Zingiberaceae) in Malaya 1. Taxonomy. – Notes Roy. Bot. Gard. Edinb. 31: 243-269.
Linhart YB. 1973. Ecological and behavioral determinants of pollen dispersal in hummingbird-pollinated Heliconia. – Amer. Nat. 107: 511-523.
Liu A-Z, Kress WJ, Wang H, Li D-Z. 2002. Insect pollination of Musella (Musaceae), a monotypic genus endemic to Yunnan, China. – Plant Syst. Evol. 235: 135-146.
Liu A-Z, Li D-Z, Li X-W. 2002. Taxonomic notes on wild bananas (Musa) from China. – Bot. Bull. Acad. Sin. 43: 77-81.
Liu A-Z, Kress WJ, Li D-Z. 2010. Phylogenetic analyses of the banana family (Musaceae) based on nuclear ribosomal (ITS) and chloroplast (trnL-F) evidence. – Taxon 59: 20-28.
Lock JM. 1984. Notes on East African Zingiberaceae. – Kew Bull. 39: 837-843.
Lock JM. 1985. Zingiberaceae. – In: Polhill RM (ed), Flora of tropical East Africa, A. A. Balkema, Rotterdam, The Netherlands, pp. 1-40.
Lock JM. 1991. A new species of Siphonochilus (Zingiberaceae) from Tanzania. – Kew Bull. 46: 269-271.
Lock JM, Hall JB, Abbiw DK. 1977. The cultivation of melegueta pepper (Aframomum melegueta) in Ghana. – Econ. Bot. 31: 321-330.
Loesener LET. 1920. Über eine neue Stromanthe-Art. – Notizbl. Bot. Gar. Mus. Berlin-Dahlem 7: 410-411.
Loesener LET. 1930a. Zingiberaceae. – In: Engler A (eds), Die natürlichen Pflanzenfamilien, 2. Aufl., Bd. 15a, W. Engelmann, Leipzig, pp. 541-640.
Loesener LET. 1930b. Marantaceae. – In: Engler A (ed), Die natürlichen Pflanzenfamilien, 2. Aufl., Bd. 15a, W. Engelmann, Leipzig, pp. 654-693.
Long H, Wen Y. 1997. Pollen morphology of Lowiaceae from China. – J. Trop. Subtrop. Bot. 5: 6-9.
Luis JG, Echeverri F, Quinines W, Brito I, Lopez M, Torres F, Cardona G, Aguiar Z, Pelaez C, Rojas M. 1993. Irenolone and emenolone: two new types of phytoalexin from Musa paradisiaca. – J. Org. Chem. 58: 4306-4308.
Luis JG, Fletcher WQ, Echeverri F, Abad T, Kishi MP, Perales A. 1995. New phenalenone-type phytoalexins from Musa acuminata (COLLA AAA) Grand Nain. – Nat. Prod. Lett. 6: 23-30.
Maas PJM. 1972. Flora Neotropica. Monograph 8. Costoideae (Zingiberaceae). – Hafner, New York.
Maas PJM. 1976. 222. Zingiberaceae. – In: Harling G, Sparre B (eds), Flora of Ecuador 6, Swedish Natural Science Research Council, Stockholm, pp. 1-49.
Maas PJM. 1977. Flora Neotropica. Monograph 18. Renealmia (Zingiberaceae-Zingiberoideae), Costoideae (Additions) (Zingiberaceae). – New York Botanical Garden, Bronx, New York.
Maas PJM. 1979. Notes on New World Zingiberaceae II. – Acta Bot. Neerl. 28: 90.
Maas PJM, Maas H. 1987. Notes on New World Zingiberaceae III. Some new species in Renealmia. – Notes Roy. Bot. Gard. Edionburgh 44: 237-248.
Maas PJM, Maas H. 1988. 223. Cannaceae. – In: Harling G, Andersson L (eds), Flora of Ecuador 32, Nord. J. Bot., Copenhagen, pp. 3-9.
Maas PJM, Maas H. 1990. Notes on New World Zingiberaceae IV. Some new species of Costus and Renealmia. – Notes Roy. Bot. Gard. Edinb. 46: 307-320.
Maas-van de Kamer H, Maas PJM. 2008. The Cannaceae of the world. – Blumea 53: 247-318.
Maas-van de Kamer H, Maas PJM, Wieringa JJ, Specht CD. 2016. Monograph of African Costaceae. – Blumea 61: 280-318.
Madulid DA. 1996.The family Zingiberaceae and the flora of the Philippines project. – In: Wu T-L, Wu Q-G, Chen Z-Y (eds), Proceedings of the 2nd symposium on the family Zingiberaceae 1-9, South China Institute of Botany, Guangzhou, People’s Republic of China.
McGahan MW. 1961. Studies on the seed of banan 1. Anatomy of the seed and embryo of Musa balbisiana. – Ann. Bot., N. S., 48: 230-238.
Mahanty HK. 1970. A cytological study of the Zingiberales with special reference to their taxonomy. – Cytologia 35: 13-49.
Malterud KD, Wollenweber E, Gomez LD. 1979. The wax of Calathea lutea (Marantaceae). – Zeitschr. Naturforsch. 34c: 157-158.
Manchester SR, Kress J. 1993. Fossil bananas (Musaceae): Ensete oregonense sp. nov. from the Eocene of western North America and its phytogeographic significance. – Amer. J. Bot. 80: 1264-1272.
Mangaly JK, Nayar J. 1990. Palynology of South Indian Zingiberaceae. – Bot. J. Linn. Soc. 103: 351-366.
Manning JC, Goldblatt P. 1989. Chromosome number in Phenacospermum and Strelitzia and the basic chromosome number in Strelitziaceae (Zingiberales). – Ann. Missouri Bot. Gard. 76: 932-933.
Mao AA, Bhaumik M. 2007. Roscoea ngainoi (Zingiberaceae) sp. nov. from Manipur, India. – Nord. J. Bot. 25: 299-302.
Martin FW. 1970. The stigmatic exudate of Strelitzia. – Phyton (Buenos Aires) 27: 47-53.
Mauritzon J. 1936. Samenbau und Embryologie einiger Scitamineen. – Acta Univ. Lund. (Lunds Univ. Årsskr.), N. F., Avd. 2, 31: 1-31.
Mello Filho LE de. 1972. Uma nova interpretação da morfologia floral de Heliconia L. f. (Musaceae). – An. Acad. Bras. Cienc. 44: 608.
Mercedes Ciciarelli M de las. 2007. Canna ascendens (Cannaceae), una nueva especie de la provincial de Buenos Aires y comentarios sobre otras especies argentines de este género. – Darwiniana 45: 188-200.
Milne-Redhead E. 1950. Notes on African Marantaceae I. Trachyphrynium as recognized by K. Schumann. – Kew Bull. 1950: 157-163.
Milne-Redhead E. 1952a. Notes on African Marantaceae II. Phrynium Willd. emend. K. Schum. in Africa. – Kew Bull. 1952: 167-170.
Milne-Redhead E. 1952b. Marantaceae. – In: Turrill WB, Milne-Redhead E (eds), Flora of tropical East Africa, The Crown Agents for the Colonies, London, pp. 1-11.
Mood J, Larsen K. 1997. Cornukaempferia, a new genus of Zingiberaceae from Thailand. –Bull. Siam Soc. Nat. Hist. 45: 217-221.
Moore HE Jr, Hyypio PA. 1970. Some comments on Strelitzia (Strelitziaceae). – Baileya 17: 65-74.
Müller F. 1885. Die Blütenpaare der Marantaceen. – Ber. Deutsch. Bot. Ges. 3: 54-56.
Müller F. 1890. Miscellen. Kreuzung von Hedychium. – Abh. Naturwiss. Ver. Bremen 11: 444.
Nakai T. 1941. Notulae ad plantas orientalis XVI. – Jap. J. Bot. 17: 189-203.
Nakai T. 1948. A new attempt in the classification of the Strelitziaceae. – Bull. Tokyo Sci. Mus. 22: 19-24.
Napp-Zinn K, Franz A. 1980. Vergleichend-anatomische Untersuchungen an petaloiden Hochblättern von Heliconia revoluta (Griggs) Standley (vorläufige Mitteilung). – Flora 170: 565-569.
Newman MF. 1990. A reconsideration of Brachychilum Peterson (Hedychieae: Zingiberaceae). – Edinburgh J. Bot. 47: 83-87.
Newman MF. 1995. Distichochlamys, a new genus from Vietnam. – Edinburgh J. Bot. 52: 65-69.
Newman SWH, Kirchoff BK. 1992. Ovary structure in the Costaceae (Zingiberales). – Intern. J. Plant Sci. 153: 471-487.
Ngamriabsakul C. 2009. Caulokaempferia sirirugsae sp. nov. (Zingiberaceae) from southern Thailand. – Nord. J. Bot. 26: 325-328.
Ngamriabsakul C, Newman MF, Cronk QCB. 2000. Phylogeny and disjunction in Roscoea (Zingiberaceae). – Edinburgh J. Bot. 57: 39-61.
Ngamriabsakul C, Newman MF, Cronk QCB. 2004. The phylogeny of the tribe Zingibereae (Zingiberaceae) based on ITS (nrDNA) and trnL-F (cpDNA) sequences. – Edinburgh J. Bot. 60: 483-507.
Nicolson DH, Steyermark JA, Sivadasan M. 1981. Thymocarpus cannoides (Marantaceae), a new genus and species from Venezuela and Brazil. – Brittonia 33: 22-24.
Nur N. 1976. Studies on pollination in Musaceae. – Ann. Bot., N. S., 40: 167-177.
Olatunji OA. 1980. The structure and development of stomata in some Zingiberales. – Notes Roy. Bot. Gard. Edinb. 38: 499-516.
Otálvaro F, Görls H, Hölscher D, Schmitt B, Echeverri F, Quiñones W, Schneider B. 2002. Dimeric phenylphenalenones from Musa acuminata and various Haemodoraceae species. – Phytochemistry 60: 61-66.
Pai RM. 1965a. Morphology of the flower in Cannaceae. – J. Biol. Sci. 8: 4-8.
Pai RM. 1965b. The floral anatomy of Elettaria cardamomum Maton. A reinvestigation. – New Phytol. 64: 187-204.
Pai RM, Tilak VD. 1965. Septal nectaries in the Scitamineae. – J. Biol. Sci. 8: 1-3.
Palla E. 1891. Ueber die Entwicklung und Bedeutung der Zellfäden im Pollen von Strelitzia reginae. – Ber. Deutsch. Bot. Ges. 9: 85-91.
Pedersen LB. 2001. Four new species of Orchidantha (Lowiaceae) from Sabah. – Nord. J. Bot. 21: 121-128.
Pedersen LB. 2004. Phylogenetic analysis of the subfamily Alpinioideae (Zingiberaceae), particularly Etlingera Giseke, based on nuclear and plastid DNA. – Plant Syst. Evol. 245: 239-258.
Pedersen LB, Johansen B. 2004. Anatomy of the unusual stigma in Orchidantha (Lowiaceae). – Amer. J. Bot. 91: 299-305.
Petersen OG. 1889a. Musaceae. – In: Engler A, Prantl K (eds), Die natürlichen Pflanzenfamilien II(6), W. Engelmann, Leipzig, pp. 1-10; Schumann K. 1897. Nachträge zu II(6), pp. 88-90.
Petersen OG. 1889b. Zingiberaceae. – In: Engler A, Prantl K (eds), Die natürlichen Pflanzenfamilien II(6), W. Engelmann, Leipzig, pp. 10-30; Schumann K. 1897. Nachträge zu II(6), pp. 90-93.
Petersen OG. 1889c. Cannaceae. – In: Engler A, Prantl K (eds), Die natürlichen Pflanzenfamilien II(6), W. Engelmann, Leipzig, pp. 30-32.
Petersen OG. 1889d. Marantaceae. – In: Engler A, Prantl K (eds), Die natürlichen Pflanzenfamilien, lI(6), W. Engelman, Leipzig, pp. 31-43; Schumann K. 1897. Nachträge zu II(6), pp. 94-96.
Pijl L van der. 1954. Xylocopa and flowers in the tropics I-III. – Proc. Kon. Ned. Akad. Wet., ser. C 57: 413-423, 541-562.
Pischtschan E, Claßen-Bockhoff R. 2008. The setting-up of tension in the style of Marantaceae. – Plant Biol. 10: 441-450.
Pischtschan E, Claßen-Bockhoff R. 2010. Anatomic insights into the thigmonastic style tissue in Marantaceae. – Plant Syst. Evol. 286: 91-102.
Pischtschan E, Ley AC, Claßen-Bockhoff R. 2010. Ontogenetic and phylogenetic diversification of the hooded staminode in Marantaceae. – Taxon 59: 1111-1125.
Poulsen AD. 1993. Two new species of Boesenbergia (Zingiberaceae) from Borneo. – Nord. J. Bot. 13: 289-294.
Poulsen AD, Leong-Škorničková J. 2017. Two new Orchidantha species (Lowiaceae) from Borneo. – Blumea 62: 157-162.
Poulsen AD, Lock JM. 1997. New species and new records of Zingiberaceae and Costaceae from tropical East Africa. – Kew Bull. 52: 601-616.
Poulsen AD, Mood J, Ibrahim H. 1999. Three new species of Etlingera (Zingiberaceae) from Borneo. – Nord. J. Bot. 19: 139-147.
Poulsen AD, Mathisen HB, Newman MF, Ardiyani M, Lofthus Ø, Bjorå CS. 2018. Sulettaria: a new ginger genus disjunct from Elettaria cardamomum. – Taxon 67: 725-738.
Prakash N, Lee YH, Goh CJ. 2000. Anther and pollen development in Heliconia. – Harvard Papers Bot. 5: 193-200.
Prince LM. 2010. Phylogenetic relationships and species delimitation in Canna (Cannaceae). – In: Seberg O, Petersen G, Barfod AS, Davis JI (eds), Diversity, phylogeny, and evolution in the monocotyledons, Aarhus University Press, Århus, pp. 307-331.
Prince LM, Kress WJ. 2006a. Phylogenetic relationships and classification in Marantaceae: insights from plastid DNA sequence data. – Taxon 55: 281-296.
Prince LM, Kress WJ. 2006b. Phylogeny and biogeography of the prayer plant family: getting to the root problem in Marantaceae. – In: Columbus JT, Friar EA, Porter JM, Prince LM, Simpson MG (eds), Monocots: comparative biology and evolution. Excluding Poales, Rancho Santa Ana Botanical Garden, Claremont, California. – Aliso 22: 645-659.
Punt W. 1968. Pollen morphology of the American species of the subfamily Costoideae (Zingiberaceae). – Rev. Palaeobot. Palynol. 7: 31-43.
Rangsiruji A. 1999. A study of the infrageneric classification of Alpinia Roxb. (Zingiberaceae) using molecular data. – Ph.D. diss., University of Edinburgh, Scotland.
Rangsiruji A, Newman MF, Cronk QCB. 2000a. Origin and relationships of Alpinia galanga (Zingiberaceae) based on molecular data. – Edinburgh J. Bot. 57: 9-37.
Rangsiruji A, Newman MF, Cronk QCB. 2000b. A study of the infrageneric classification of Alpinia (Zingiberaceae) based on the ITS region of nuclear rDNA and the trnL-F spacer of chloroplast DNA- In: Wilson KL, Morrison DA (eds), Monocots: systematics and evolution, CSIRO Publ, Collingwood, Australia, pp. 695-709.
Raninandran PN, Babu KN (eds). 2005. Ginger: the genus Zingiber. – CRC Press, Boca Raton.
Rao AS, Verma DM. 1971. Curcumorpha – a new genus of Zingiberaceae. – Bull. Bot. Survey India 13: 339-341.
Rao VS. 1963. The epigynous glands of Zingiberaceae. – New Phytol. 62: 342-349.
Rao VS, Karnik H, Gupte K. 1954. The floral anatomy of some Scitamineae I. – J. Indian Bot. Soc. 33: 118-147.
Rehse T, Kress WJ. 2003. Distichochlamys rubrostriata (Zingiberaceae), a new species from northern Vietnam. – Brittonia 55: 205-208.
Ren P-Y, Liu M, Li Q-J. 2007. An example of flexistyly in a wild cardamom species (Amomum maximum (Zingiberaceae)). – Plant Syst. Evol. 267: 147-154.
Reynolds PK. 1927. The banana. – Houghton-Mifflin Co., Riverside Press, Cambridge.
Ridley HN. 1887. Angolan Scitamineae. – J. Bot. 25: 129-135.
Ridley HN. 1899. The Scitamineae of the Malay Peninsula . – J. Straits Branch Roy. Asiat. Soc. 32: 85-184.
Ridley HN. 1909. The Scitamineae of the Philippines. – Philipp. J. Sci. 4: 155-193.
Rodríguez G. 1954a. Sobre Heliconia latispatha y una espécie no descrita confundida con ella. – Acta Biol. Venezuelica 1: 209-212.
Rodríguez G. 1954b. Revisión del género Heliconia en Venezuela. – Bot. Soc. Venezolana Ci. Nat. 15: 117-130.
Rodríguez-de la Rosa RA, Cevallos-Ferriz SRS. 1994. Upper Cretaceous zingiberalean fruits with in situ seeds from southeastern Coahuila, Mexico. – Intern. J. Plant Sci. 155: 786-805.
Rose M-J, Barthlott W. 1995. Pollen-connecting threads in Heliconia (Heliconiaceae). – Plant Syst. Evol. 195: 61-65.
Ross EM. 1987. Musaceae. – In: George AS (ed), Flora of Australia 45, Australian Government Publ. Service, Canberra, pp. 16-19.
Rowley JR, Skvarla JJ. 1975. The glycocalyx and initiation of exine spinules on microspores of Canna. – Amer. J. Bot. 62: 479-485.
Rowley JR, Skvarla JJ. 1986. Development of the pollen grain wall in Canna. – Nord. J. Bot. 6: 39-65.
Saad SI, Ibrahim RK. 1965. Palynological and biochemical studies of Scitamineae. – J. Palynol. 1: 62-66.
Sabu M. 2006. Zingiberaceae and Costaceae of south India. – Ind. Assoc. Angiosperm Taxonomy, Dept. of Bot., Calicut Univ., pp. 225-250.
Saensouk P, Chantaranothai P, Theerakulpisut P. 2009. Pollen morphology of the genus Cornukaempferia (Zingiberaceae) in Thailand. – J. Syst. Evol. 47: 139-143.
Saensouk S. 2006. Taxonomy and biology of the genus Alpinia Roxb. (Zingiberaceae) in Thailand. – Ph.D. diss., Khon Kaen University, Thailand.
Sagot P. 1887. Sur le genre bananier. – Bull. Soc. Bot. France 34: 328-330.
Sakai S, Inoue T. 1999. A new pollination system: dung beetle pollination discovered in Orchidanthera inouei (Lowiaceae, Zingiberales) in Sarawak, Malaysia. – Amer. J. Bot. 86: 56-61.
Sakai S, Nagamasu H. 1998. Systematic studies of Bornean Zingiberaceae I. Amomum in Lambir Hills, Sarawak. – Edinburgh J. Bot. 55: 45-64.
Sakai S, Nagamasu H. 2000. Systematic studies of Bornean Zingiberaceae III. Tamijia: a new genus. – Edinburgh J. Bot. 57: 245-255.
Salzman S, Driscoll HE, Renner T, André T, Shen S, Specht CD. 2015. Spiraling into history: a molecular phylogeny and investigation of biogeographic origins and floral evolution for the genus Costus. – Syst. Bot. 40: 104-115.
Sam YY, Takano A, Ibrahim H, Záveská E, Aziz F. 2016. Borneocola (Zingiberaceae), a new genus from Borneo. – PhytoKeys 75: 31-55.
Sardos J, Perrier X, Doležel, Hřibová E, Christelová P, Van den Houwe I, Kilian A, Roux N. 2016. DArT whole genome profiling provides insights on the evolution and taxonomy of edible Banana (Musa spp.). – Ann. Bot. 118: 1269-1278.
Särkinen TE, Newman MF, Maas PJM, Maas H, Poulsen AD, Harris DJ, Richardson JE, Clark A, Hollingsworth M, Pennington RT. 2007. Recent oceanic long-distance dispersal and divergence in the amphi-Atlantic rain forest genus Renealmia L. f. (Zingiberaceae). – Mol. Phylogen. Evol. 44: 968-980.
Sato D. 1960. The karyotype analysis in Zingiberales with special reference to the proto-karyotype and stable karyotype. – Sci. Pap. Coll. Gen. Educ. Univ. Tokyo 10: 225-243.
Schachner J. 1924. Beiträge zur Kenntnis der Blüten- und Samenentwicklung der Scitamineen. – Flora 117: 16-40.
Schemske DW. 1980. The evolutionary significance of extrafloral nectar production by Costus woodsonii (Zingiberaceae): an experimental analysis of ant protection. – J. Ecol. 68: 959-967.
Schumann K. 1892 [1893]. Zingiberaceae africanae. – Engl. Bot. Jahrb. Syst. 15: 419-424.
Schumann K. 1899. Monographie der Zingiberaceae von Malesien und Papuasien. – Engl. Bot. Jahrb. Syst. 27: 259-350.
Scott-Elliot GF. 1980. Note on the fertilization of Musa, Strelitzia reginae, and Ravenala madagascariensis. – Ann. Bot., N. S., 45: 259-263.
Scutch AF. 1932. Anatomy of the axis of the banana. – Bot. Gaz. 93: 233-258.
Searle RJ, Hedderson TAJ. 2000. A preliminary phylogeny of the Hedychieae tribe (Zingiberaceae) based on ITS sequences of the nuclear rRNA cistron. – In: Wilson KL, Morrison DA (eds), Monocots: systematics and evolution, CSIRO Publ., Melbourne, pp. 710-718.
Segeren W, Maas PJM. 1971. The genus Canna in northern South America. – Acta Bot. Neerl. 20: 663-680.
Sharma AK, Bhattacharyya NK. 1958. Inconstancy in chromosome complements in species of Maranta and Calathea. – Proc. Natl. Inst. Sci. India, Sect. B, 24: 101-117.
Sharma AK, Bhattacharyya NK. 1959. Cytology of several members of Zingiberaceae and a study of the inconstancy of their chromosome complements. – La Cellule 59: 299-346.
Shepherd K. 1990. Observations on Musa taxonomy. – In: Jerret RL (ed), Identification of genetic diversity in the genus Musa, INIBAP, Montpellier, pp. 158-165.
Simão DG, Scatena VL, Bouman F. 2006. Developmental anatomy and morphology of the ovule and seed of Heliconia (Heliconiaceae, Zingiberales). – Plant Biol. 8: 143-154.
Simão DG, Scatena VL, Bouman F. 2007. Anther development, microsporogenesis and microgametogenesis in Heliconia (Heliconiaceae, Zingiberales). – Flora 202: 148-160.
Simmonds NW. 1953. The development of the banana fruit. – J. Exp. Bot. 4: 87-105.
Simmonds NW. 1959. Bananas. – Longman, Green & Co., London.
Simmonds NW. 1960a. Megasporogenesis and female fertility in three edible triploid bananas. – J. Genet. 57: 269-278.
Simmonds NW. 1960b. Notes on banana taxonomy. – Kew Bull. 14: 198-212.
Simmonds NW. 1962. The evolution of the bananas. – Longman, Green & Co., London.
Simmonds NW. 1966. Bananas, 2nd ed. – Longmans, Green & Co., London.
Simmonds NW, Weatherup STC. 1990. Numerical taxonomy of the wild bananas (Musa). – New Phytol. 115: 567-571.
Sirirugsa P. 1987. Three new species and one new combination in Boesenbergia (Zingiberaceae) from Thailand. – Nord. J. Bot. 7: 421-425.
Sirirugsa P. 1989. The genus Kaempferia (Zingiberaceae) in Thailand. – Nord. J. Bot. 9: 257-260.
Sirirugsa P. 1992. A revision of the genus Boesenbergia Kuntz (Zingiberaceae) in Thailand. – Nat. Hist. Bull. Siam Soc. 40: 67-90.
Sirirugsa P, Larsen K. 1995. The genus Hedychium (Zingiberaceae) in Thailand. – Nord. J. Bot. 15: 301-304.
Sivori E, Nakayama F, Cigliano E. 1968. Germination of achira seed (Canna sp.) approximately 550 years old. – Nature 219: 1269-1270.
Skutch AF. 1930. Unrolling of leaves of Musa sapientium and some related plants and their reactions to environmental aridity. – Bot. Gaz. 90: 337-365.
Skvarla J, Rowley JR. 1970. The pollen wall of Canna and its similarity to the germinal apertures of other pollen. – Amer. J. Bot. 57: 519-529.
Smith JF, Kress WJ, Zimmer EA. 1993. Phylogenetic analysis of the Zingiberales based on rbcL sequences. – Ann. Missouri Bot. Gard. 80: 620-630.
Smith RM. 1972. The genus Burbidgea. – Notes Roy. Bot. Gard. Edinb. 31: 297-306.
Smith RM. 1980. A new genus of Zingiberaceae from N. Burma. – Notes Roy. Bot. Gard. Edinb. 38: 13-17.
Smith RM. 1981. Zingiberaceae. Synoptic keys to the tribes Zingibereae, Globbeae, Hedychieae, Alpinieae (in part). – Royal Botanic Gardens, Edinburgh.
Smith RM. 1982. Systematic notes on, and new species of Zingiberaceae of the Gunung Mulu National Park. – Bot. J. Linn. Soc. 85: 36-73.
Smith RM. 1985. A review of Bornean Zingiberaceae I (Alpinieae p. p.). – Notes Roy. Bot. Gard. Edinb. 42: 261-314.
Smith RM. 1987a. Zingiberaceae. – In: George AS (ed), Flora of Australia 45, Australian Government Publ. Service, Canberra, pp. 19-34.
Smith RM. 1987b. Costaceae. – In: George AS (ed), Flora of Australia 45, Australian Government Publ. Service, Canberra, pp. 34-36.
Smith RM. 1987c. A review of the Bornean Zingiberaceae III. (Hedychieae). – Notes Roy. Bot. Gard. Edinb. 44: 203-232.
Smith RM. 1988. A review of Bornean Zingiberaceae IV. (Globbeae). – Notes Roy. Bot. Gard. Edinb. 45: 1-19.
Smith RM. 1989. A review of Bornean Zingiberaceae V. (Zingiber). – Notes Roy. Bot. Gard. Edinb. 45: 409-423.
Smith RM. 1990a. Alpinia (Zingiberaceae): a proposed new infrageneric classification. – Edinburgh J. Bot. 47: 1-75.
Smith RM. 1990b. Psychanthus (K. Schum.) Ridley (Zingiberaceae): its acceptance at generic level. – Edinburgh J. Bot. 47: 77-82.
Song J-J, Liao J-P, Tang Y-J, Chen Z-Y. 2004. Chromosome numbers in Orchidanthera (Lowiaceae) and their biogeographic and systematic implications. – Ann. Bot. Fenn. 41: 429-433.
Spearing JK. 1977. A note on closed leaf sheaths in Zingibereae-Zingiberoideae. – Notes Roy. Bot. Gard. Edinb. 35: 217-220.
Specht CD. 2005. Phylogenetics, floral evolution and rapid radiation in the tropical monocot family Costaceae (Zingiberales). – In: Sharma AK, Sharma A (eds), Plant genome: biodiversity and evolution. Vol. 1, part B. Phanerogams (higher groups), Scientific Publ., Enfield, New Hampshire, pp. 29-60.
Specht CD. 2006a. Systematics and evolution of the tropical monocot family Costaceae (Zingiberales): a multiple dataset approach. – Syst. Bot. 31: 89-106.
Specht CD. 2006b. Gondwanan vicariance or dispersal in the tropics? The biogeographic history of the tropical monocot family Costaceae (Zingiberales). – In: Columbus JT, Friar EA, Porter JM, Prince LM, Simpson MG (eds), Monocots: comparative biology and evolution. Excluding Poales, Rancho Santa Ana Botanical Garden, Claremont, California. – Aliso 22: 633-644.
Specht CD, Stevenson DW. 2006. A new phylogeny-based generic taxonomy for the monocot family Costaceae (Zingiberales). – Taxon 555: 153-163.
Specht CD, Kress WJ, Stevenson DW, DeSalle R. 2001. A molecular phylogeny of Costaceae (Zingiberales). – Mol. Phylogen. Evol. 21: 333-345. – Errata: Mol. Phylogen. Evol. 23 (2002): 95.
Standley PC. 1937. Flora of Costa Rica 1. Marantaceae. – Publ. Field Mus. Nat. Hist. 18: 191-196.
Standley PC, Steyermark JA. 1952. Flora of Guatemala: Marantaceae. – Fieldiana Bot. 24: 207-221.
Stauffer FW, Barfod A, Endress PK. 2009. Floral structure in Licuala peltata (Arecaceae: Coryphoideae), with special reference to the architecture of the unusual labyrinthine nectary. – Bot. J. Linn. Soc. 161: 66-77.
Stiles FG. 1975. Ecology, flowering phenology, and hummingbird pollination of some Costa Rican Heliconia species. – Ecology 56: 285-301.
Stiles FG. 1979. Notes on the natural history of Heliconia (Musaceae) in Costa Rica. – Brenesia 15 [Suppl.]: 151-180.
Stone DE. 1987. Developmental evidence for the convergence of Sassafras (Laurales) and Heliconia (Zingiberales) pollen. – Grana 26: 179-191.
Stone DE, Sellers SC, Kress WJ. 1979. Ontogeny of exineless pollen in Heliconia, a banana relative. – Ann. Missouri Bot. Gard. 66: 701-730.
Stone DE, Sellers SC, Kress WJ. 1981. Ontogenetic and evolutionary implications of a neotenous exine in Tapeinochilos (Zingiberales: Costaceae) pollen. – Amer. J. Bot. 68: 49-63.
Suksathan P, Borchsenius F. 2003. Two new species of Stachyphrynium (Marantaceae) from SE Asia. – Willdenowia 33: 403-408.
Suksathan P, Borchsenius F. 2005. Nomenclatural synopsis of Marantaceae in Thailand. – Taxon 54: 1083-1090.
Takano A, Nagamasu H. 2007. Myxochlamys (Zingiberaceae), a new genus from Borneo. – Acta Phytotax. Geobot. 58: 19-32.
Takano A, Okada H. 2002. Multiple occurrences of triploid formation in Globba (Zingiberaceae) from molecular evidence. – Plant Syst. Evol. 230: 143-159.
Takano A, Okada H. 2003. Taxonomy of Globba (Zingiberaceae) in Sumatra, Indonesia. – Syst. Bot. 28: 524-546.
Tanaka N. 2001. Taxonomic revision of the family Cannaceae in the New World and Asia. – Makinoa, n. s. 1: 1-74.
Tanaka N. 2008. A new species of the genus Canna (Cannaceae) from Eastern Honduras. – J. Jap. Bot. 88: 7/10.
Tanaka N, Uchiyama H, Matoba H, Koyama T. 2009. Karyological analysis of the genus Canna (Cannaceae). – Plant Syst. Evol. 280: 45-51.
Tang Y-J, Xie Z-Y, Liao J-P, Wu Q-G. 2005. Anatomy and histochemistry of the seeds in ginger families of Zingiberales and their systematic significance. – Acta Bot. Bor.-Occid. Sin. 25: 343-354. [In Chinese]
Theerakulpisut P, Triboun P, Mahakham W, Maensiri D, Khampila J, Chantaranothai P. 2012. Phylogeny of the genus Zingiber (Zingiberaceae) based on nuclear ITS sequence data. – Kew Bull. 67: 389-395.
Theilade I. 1999. A synopsis of the genus Zingiber (Zingiberaceae) in Thailand. – Nord. J. Bot. 19: 389-410.
Theilade I, Mood J. 1997. Five new species of Zingiber (Zingiberaceae) from Borneo. – Nord. J. Bot. 17: 337-347.
Theilade I, Mærsk-Møller ML, Theilade J, Larsen K. 1993. Pollen morphology and structure of Zingiber (Zingiberaceae). – Grana 32: 338-342.
Thompson J Mclean. 1933. Studies in advancing sterility VI. The theory of scitaminean flowering. – Publ. Hartley Bot. Labor. 11: 1-144.
Tilak VD. 1968. On the morphology of the perianth in Musa L. – Marathwada Univ. J. 7: 7-10.
Tilak VD, Pai RM. 1966. Studies in the floral morphology of the Marantaceae I. Vascular anatomy of the flower of Schumannianthus virgatus Rolfe, with special reference to the labellum. – Can. J. Bot. 44: 1365-1370.
Tilak VD, Pai RM. 1968. Studies in the floral morphology of the Marantaceae II. Vascular anatomy of the genus Phrynium Willd. – Proc. Indian Acad. Sci., Sect. B, 68: 240-249.
Tilak VD, Pai RM. 1970. Studies in the floral morphology of the Marantaceae III. Vascular anatomy of some species of the genus Calathea. – Marathwada Univ. J. 9, Sci. 2: 31-41.
Tilak VD, Pai RM. 1974. The floral anatomy of Ensete superbum (Roxb.) Cheesem. – Proc. Indian Acad. Sci., Sect. B, 80: 253-261.
Tomlinson PB. 1956. Studies in the systematic anatomy of the Zingiberaceae. – Bot. J. Linn. Soc. 55: 547-592.
Tomlinson PB. 1959. An anatomical approach to the classification of the Musaceae. – Bot. J. Linn. Soc. 55: 779-809.
Tomlinson PB. 1960. The anatomy of Phenakospermum (Musaceae). – J. Arnold Arbor. 41: 287-297.
Tomlinson PB. 1961a. The anatomy of Canna. – Bot. J. Linn. Soc. 56: 467-473.
Tomlinson PB. 1961b. Morphological and anatomical characteristics of the Marantaceae. – Bot. J. Linn. Soc. 58: 55-78.
Tomlinson PB. 1962. Phylogeny of the Scitamineae – morphological and anatomical considerations. – Evolution 16: 192-213.
Tomlinson PB. 1969. Classification of the Zingiberales (Scitamineae) with special reference to anatomical evidence. – In: Metcalfe CR (ed), Anatomy of the monocotyledons 3, Oxford, pp. 295-302.
Tong SQ. 1985. Paramomum, a new genus of Zingiberaceae from Yunnan. – Acta Bot. Yunnan. 7: 309-312.
Tran VN. 1974. Sur le callus des marantacées. – Bull. Soc. Bot. France 121: 97-108.
Triboun P. 2006. Biogeography and biodiversity of the genus Zingiber in Thailand. – PhD diss., Khon Kaen University, Thailand, pp. 140-144.
Trivedi BS, Verma CL. 1970. Silicified pseudostem of Canna L. from early Eocene of Deccan Intertrappean beds, M. P., India. – Curr. Sci. 39: 442-443.
Trivedi BS, Verma CL. 1971. The structure of pseudostem and root of Cannaites intertrappea gen. et sp. nov. from the Deccan Intertrappean beds of Madhya Pradesh, India. – Palaeontographica, Abt. B, 132: 175-185.
Trivedi BS, Verma CL. 1972. Occurrence of Heliconiaites mohgaonensis gen. et sp. nov. from the Early Eocene of Deccan Intertrappean series, M. P., India. – Palaeontographica, Abt. B, 139: 73-82.
Ude G, Pillay M, Nwakanma D, Tenkouano A. 2002. Analysis of genetic diversity and sectional relationships in Musa using AFLP markers. – Theor. Appl. Genet. 104: 1239-1245.
Uma E, Muthukumar T. 2014. Comparative root
morphological anatomy of Zingiberaceae. – Syst. Biodivers.
12: 195-209.
Valeton T. 1918. New notes on the Zingiberaceae of Java and Malaya. – Bull. Jard. Bot. Buitenzorg, sér. II, 27: 1-166.
Varela de Vega A. 1972. Interpretación morfológica de las flores masculinas de algunas espécies y variedades de Musa. – Rev. Fac. Agron. Maracay 4(3): 65-78.
Venkatasubban KR. 1946. A preliminary survey of chromosome numbers in Scitaminae of Bentham and Hooker. – Proc. Indian Acad. Sci., Sect. B, 23(b): 281-300.
Venkataswaralu V. 1937. A note on the development of the embryo sac in Phrynium capitatum W. – J. Indian Bot. Soc. 16: 95-98.
Venter HA van de. 1976a. Microsporogenesis and -gametogenesis in Strelitzia reginae Ait. – J. South Afr. Bot. 42: 25-31.
Venter HA van de. 1976b. A note on ovular outgrowths in the species of Strelitzia Ait. – J. South Afr. Bot. 42: 37-39.
Venter HA van de. 1976c. Notes on the morphology of the embryo and seedling of Strelitzia reginae Ait. – J. South Afr. Bot. 42: 63-69.
Vieira S, Souza VC. 2008. Four new species of Maranta L. (Marantaceae) from Brazil. – Bot. J. Linn. Soc. 158: 131-139.
Vieira S, Maas PJM, Borchsenius F. 2012. Taxonomic revision of Myrosma (Marantaceae). – Blumea 57: 125-130.
Wagner A. 1894. Zur Anatomie und Biologie der Blüthe von Strelitzia reginae. – Ber. Deutsch. Bot. Ges. 12: 53-72.
Weber A. 1991. Globba unifolia and G. corneri stat. et nom. nov. (Zingiberaceae; Peninsular Malaysia). – Plant Syst. Evol. 174: 1-4.
Weber H. 1958. Die Wurzelverdickungen von Calathea macrosepala Schum. und einigen anderen monokotylen Pflanzen. – Beitr. Biol. Pflanzen 34: 177-193.
Weisse A. 1932. Zur Kenntnis der Blattstellungsverhältnisse bei den Zingiberaceen. – Ber. Deutsch. Bot. Ges. 50A: 327-366.
Weisse A. 1933. Die Art der Distichie an den Achsensprossen von Zingiber. – Ber. Deutsch. Bot. Ges. 51: 13-20.
Wen Y, Liao J, Wu Q. 1997. Anatomy and histochemistry of the seed of Orchidanthera sinensis T. L. Wu. – Guihaia 17: 235-241. [In Chinese]
West JP, Cowley J. 1993. Floristic notes and chromosome numbers of some Chinese Roscoea (Zingiberaceae). – Kew Bull. 48: 799-803.
Williams CA, Harborne JB. 1977. The leaf flavonoids of the Zingiberales. – Biochem. Syst. Ecol. 5: 221-229.
Williams KJ, Kress WJ, Thet Htun. 2003. A striking new epiphytic Hedychium (Zingiberaceae) from Myanmar with a discussion on some anomalous related genera. – Edinburgh J. Bot. 60: 43-48.
Williams KJ, Kress WJ, Manos PS. 2004. The phylogeny, evolution and classification of the genus Globba (Zingiberaceae): appendages do matter. – Amer. J. Bot. 91: 100-114.
Winkler H. 1908. Neue Kameruner Phanerogamen aus verschiedenen Familien. Zingiberaceae. – Engl. Bot. Jahrb. Syst. 41: 275.
Winkler H. 1930a. Musaceae. – In: Engler A (ed), Die natürlichen Pflanzenfamilien, 2. Aufl., Bd. 15a, W. Engelmann, Leipzig, pp. 505-541.
Winkler H. 1930b. Cannaceae. – In: Engler A (ed), Die natürlichen Pflanzenfamilien, 2. Aufl., Bd. 15a, W. Engelmann, Leipzig, pp. 640-654.
Wong C, Kiew R, Agent G, Set O, Lee SK, Gan YY. 2002. Assessment of validity of sections in Musa (Musaceae) using AFLP. – Ann. Bot. 90: 231-238.
Wood TH, Whitten WM, Williams NH. 2000. Phylogeny of Hedychium and related genera (Zingiberaceae) based on ITS sequence data. – Edinburgh J. Bot. 57: 261-270.
Woodson RE, Schery RW. 1945. Flora of Panama. Marantaceae. – Ann. Missouri Bot. Gard. 32: 81-105.
Wu F. 1964. Lowiaceae, a family new to the flora of China. – Acta Phytotaxon. Sin. 9: 335-343.
Wu T-L. 1997. Notes on Lowiaceae, Musaceae, and Zingiberaceae for the Flora of China. – Novon 7: 440-442.
Wu T-L, Wu Q-G, Chen Z-Y. 1996. Proceedings of the second symposium on the family Zingiberaceae. – Zhongshan University Press.
Xia Y-M, Kress WJ, Prince LM. 2004. Phylogenetic analyses of Amomum (Alpinioideae: Zingiberaceae) using ITS and matK DNA sequence data. – Syst. Bot. 29: 334-344.
Xue CY, Wang H, Li D-Z. 2007. Female gametophyte and seed development in Musella lasiocarpa (Musaceae), a monotypic genus endemic to southwestern China. – Can. J. Bot. 85: 964-975.
Záveská E, Fér T, Šída O, Krak K, Marhold K, Leong-Škorničková J. 2012. Phylogeny of Curcuma (Zingiberaceae) based on plastid and nuclear sequences: proposal of the new subgenus Ecomata. – Taxon 61: 747-763.
Zhao J-L, Zhong J, Fan Y-L, Xia Y-M, Li Q-J. 2017. A preliminary species-level phylogeny of the alpine ginger Roscoea: implications for speciation. – J. Syst. Evol. 55: 215-224.