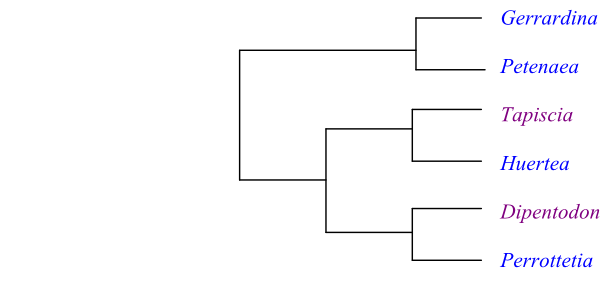
Cladogram of Huerteales based on DNA sequence data (Christenhusz & al. 2010)
[Huerteales+[Capparales+Malvales]]
Habit Bisexual or dioecious, evergreen or deciduous trees or shrubs (rarely lianas).
Vegetative anatomy Phellogen? Vessel elements with usually scalariform (rarely simple) perforation plates; lateral pits scalariform, opposite or alternate, simple or bordered pits. Imperforate tracheary xylem elements fibre tracheids or libriform fibres (rarely tracheids) with simple or bordered pits, septate or non-septate. Wood rays uniseriate or multiseriate, heterocellular. Axial parenchyma paratracheal diffuse or absent. Sieve tube plastids? Nodes 3:3, 4:4 or 5:5, tri-, tetra- or pentalacunar with three to five leaf traces, respectively. Sclereids at least sometimes present. Prismatic calciumoxalate crystals often abundant.
Trichomes Hairs unicellular or multicellular, uniseriate, simple or branched, or absent.
Leaves Alternate (spiral or distichous), simple, entire, with ? ptyxis. Stipules cauline, early caducous; leaf sheath absent. Petiole vascular bundle transection arcuate or annular. Venation pinnate, semicraspedodromous, or palmate. Stomata usually anomocytic. Cuticular wax crystalloids? Usually with domatia in abaxial vein axils. Epidermis sometimes with mucilaginous idioblasts. Leaf margin serrate, biserrate (sometimes glandular-serrate) or entire; leaf teeth sometimes cunonioid.
Inflorescence Terminal or axillary, cymose panicle, thyrsoid, spherical, umbel-, raceme- or spike-like.
Flowers Actinomorphic, small. Hypanthium present. Hypogyny. Sepals (four or) five (to seven), with valvate, imbricate or open aestivation, free or more or less connate. Petals (four or) five (to seven), with valvate or imbricate quincuncial aestivation, connate at base or free (sometimes absent). Nectaries staminodial, antepetalous, or nectariferous disc intrastaminal, annular, entire or lobed.
Androecium Stamens (four or) five (to twelve), antesepalous, alternipetalous or antepetalous. Filaments free from each other and from tepals, or connate into tubular, infundibuliform, cup-shaped or spherical structure, free from tepals. Anthers basifixed or dorsifixed, non-versatile, tetrasporangiate, introrse or latrorse, longicidal (dehiscing by longitudinal slits, or apical slit opening along anther). Tapetum secretory? Staminodia as five (to seven) nectariferous glands, or absent.
Pollen grains Microsporogenesis simultaneous? Pollen grains tricolpor(oid)ate, shed as monads, ?-celled at dispersal. Exine tectate or semitectate, with columellate infratectum, perforate, microreticulate, reticulate or finely reticulate.
Gynoecium Pistil composed of two to five connate carpels (septa sometimes incomplete apically). Ovary superior, unilocular (sometimes divided by apical or basal septum). Style usually single, simple (sometimes bifid). Stigma punctate, subcapitate or somewhat bilobate, type? Pistillodium absent.
Ovules Placentation free central, axile, basal-axile, basal-lateral, or apical. Ovules one to numerous per carpel (rarely one per ovary), anatropous, pendulous or erect, bitegmic, crassinucellar. Micropyle endostomal? Megagametophyte monosporous, Polygonum type? Endosperm development nuclear? Endosperm haustoria? Embryogenesis?
Fruit A drupaceous septicidal? capsule, a drupe or a berry.
Seeds Aril absent or present. Testa with palisade tissue inside collapsed polygonal cells, or with sarcotesta, often with strongly bullate chalaza. Exotestal cells prismatic (malpighian cells), unevenly prolonged. Endotesta with rod-like structures on inner tangential cell walls. Exotegmen palisade, often fibrous (with laterally compressed fibres), with malpighian cells with lignified walls. Endotegmen? Perisperm not developed. Endosperm usually copious, starchy. Embryo small or medium-sized, straight, chlorophyll? Cotyledons two. Germination phanerocotylar?
Cytology n = 10, 15, 17
DNA Plastid gene infA lost/defunct? Mitochondrial intron coxII.i3 lost?
Phytochemistry 4,5-dihydroblumenol A (a norisoprenoid) present. Cyanogenic glycosides not found.
Systematics Huerteales are sister-group to the clade [Capparales+Malvales]. A probable topology is [[Gerrardinaceae+Petenaeaceae]+[Tapisciaceae+Dipentodontaceae]]. According to Stevens (2001 onwards) a potential synapomorphy for the clade [Tapisciaceae+Dipentodontaceae] may be absence of wood parenchyma.
|
|
Cladogram of Huerteales based on DNA sequence data (Christenhusz & al. 2010) |
DIPENTODONTACEAE Merr. |
Dipentodontales C. Y. Wu in Acta Phytotaxon. Sin. 40: 305. 2002
Genera/species 2/c 20
Distribution Northeastern India, eastern Himalayas, northern Burma, southeastern Tibet, southwestern and southern China, Taiwan, Vietnam, Malesia to New Guinea, northeastern Queensland, the Hawaiian Islands, Mexico, Central America, the northern Andes south to Peru.
Fossils Unknown.
Habit Bisexual or dioecious, evergreen trees or shrubs.
Vegetative anatomy Phellogen? Vessel elements with usually scalariform (in Perrottetia sometimes simple) perforation plates; lateral pits scalariform, opposite or alternate, simple or bordered pits. Imperforate tracheary xylem elements libriform fibres or fibre tracheids (fibre tracheids absent in Perrottetia) with simple or bordered pits, septate (Dipentodon) or non-septate (Perrottetia). Wood rays multiseriate, heterocellular. Axial parenchyma absent (Dipentodon) or paratracheal diffuse (Perrottetia). Sieve tube plastids ? type. Nodes? Sclereids present (Perrottetia). Prismatic calciumoxalate crystals abundant.
Trichomes Hairs unicellular, uniseriate, simple, or absent.
Leaves Alternate (distichous or spiral), simple, entire, with ? ptyxis. Stipules cauline, large, lobed, early caducous; leaf sheath absent. Petiole vascular bundle transection arcuate (Dipentodon) or compressed annular (Perrottetia); petiole with wing bundles and adaxial inverted bundle (Perrottetia). Venation pinnate. Stomata usually anomocytic (sometimes anisocytic). Cuticular wax crystalloids? Epidermis in Perrottetia with mucilage cells (absent in Dipentodon). Usually with abaxial domatia (pits) in vein axils. Leaf margin serrate (sometimes glandular serrate) or entire.
Inflorescence Axillary, thyrsoid (Perrottetia) or spherical, umbel-like (pseudo-umbel), c. 25- to c. 30-flowered, with involucrum consisting of four or five small caducous bracts (Dipentodon).
Flowers Actinomorphic, small. Hypanthium short and wide (Dipentodon). Hypogyny (Dipentodon) or half epigyny (Perrottetia). Sepals (four or) five (to eight), with valvate (Dipentodon) or open to valvate or slightly imbricate (Perrottetia) aestivation, connate at base (Dipentodon) or free (Perrottetia), persistent, in Perrottetia containing idioblasts with thickened inner tangential cell walls. Petals (four or) five (to seven), with valvate or slightly imbricate aestivation, sepaloid, connate at base (Dipentodon) or free (Perrottetia), persistent. Nectaries staminodial, antepetalous (Dipentodon) or nectariferous disc intrastaminal, annular, entire or lobate.
Androecium Stamens (four or) five (to eight), haplostemonous, antesepalous, alternipetalous. Filaments free from each other and from tepals. Anthers basifixed, non-versatile, tetrasporangiate, introrse, longicidal (dehiscing by longitudinal slits). Tapetum secretory? Staminodia? as (four or) five (to seven) nectariferous glands alternating with stamens.
Pollen grains Microsporogenesis simultaneous? Pollen grains tricolporate, shed as monads, ?-celled at dispersal. Exine?, with ? infratectum, sculpturing?
Gynoecium Pistil composed of three or four (Dipentodon) or two (Perrottetia) connate carpels (septa in Dipentodon incomplete, apical). Ovary superior (Dipentodon) or semi-inferior (Perrottetia), unilocular (Dipentodon) or bilocular (Perrottetia, with apical septum). Style single, entire. Stigma punctate (Dipentodon) or subcapitate to indistinctly bilobate (Perrottetia), type? Pistillodium absent.
Ovules Placentation largely free central, axile at base (Dipentodon) or basal-lateral (Perrottetia). Ovules one per ovary (Dipentodon) or two per carpel (Perrottetia), orthotropous, apotropous to epitropous, bitegmic?, crassinucellar? Micropyle endostomal (Perrottetia). Outer integument ? cell layers thick. Inner integument ? cell layers thick. Megagametophyte monosporous, Polygonum type? Endosperm development nuclear? Endospermhaustoria? Embryogenesis?
Fruit A single-seeded drupaceous septicidal? capsule, dehiscing from base upwards (Dipentodon), or a two- to four-seeded dry berry (Perrottetia) with persistent calyx and corolla.
Seeds Aril absent. Testa with palisade tissue with obliquely orientated ribbon cells and inside these collapsed polygonal cells (Dipentodon), or with thin-walled, more or less fleshy sarcotesta, with exotestal cells prismatic (malpighian cells), unevenly prolonged and endotesta with rod-like structures on inner tangential cell walls (Perrottetia). Exotegmen palisade, furrowed, with malpighian cells with lignified walls (Perrottetia). Endotegmen? Perisperm not developed. Endosperm starchy (Perrottetia). Embryo small or medium-sized, straight (Dipentodon), chlorophyll? Cotyledons two. Germination phanerocotylar?
Cytology n = 17 (Dipentodon), n = 10 (Perrottetia)
DNA
Phytochemistry Very insufficiently known. 4,5-dihydroblumenol A (a norisoprenoid) present in Perrottetia.
Use Unknown.
Systematics Dipentodon (1; D. sinicus; northeastern India, eastern Himalayas, northern Burma, southeastern Tibet, southwestern China, Vietnam), Perrottetia (c 20; southern China, Taiwan, Malesia to New Guinea, northeastern Queensland, the Hawaiian Islands, Mexico, Central America, the northern Andes to Peru).
Perrottetia is sister to Dipentodon, according to Zhang & Simmons (2006), Worberg & al. (2009), and Christenhusz & al. (2010), and Dipentodontaceae are sister-group to Tapisciaceae.
GERRARDINACEAE M. H. Alford |
Genera/species 1/2
Distribution Tropical East Africa, southernmost and southeasternmost Africa.
Fossils Unknown.
Habit Bisexual, deciduous? shrubs or small trees (sometimes lianas).
Vegetative anatomy Phellogen? Vessel elements with scalariform perforation plates; lateral pits almost absent (scalariform and/or opposite when present), bordered pits. Imperforate tracheary xylem elements fibre tracheids (sometimes also tracheids) with bordered pits, usually non-septate? Wood rays uniseriate, biseriate or multiseriate, heterocellular. Axial parenchyma absent? Sieve tube plastids ? type. Nodes? Crystals?
Trichomes Hairs?
Leaves Alternate (spiral), simple, entire, with ? ptyxis. Stipules present (cauline?); leaf sheath absent. Petiole bundle transection arcuate, or petiole with three vascular bundles in one common bow above thick-walled fibres. Venation pinnate, festoon-shaped semicraspedodromous. Stomata? Cuticular wax crystalloids? Mesophyll with calciumoxalate mainly as druses. Epidermis with mucilage cells. Leaf margin serrate or biserrate; leaf teeth gland-tipped, cunonioid (lateral veins proceeding into tooth, with one branch to vein above).
Inflorescence Axillary, cymose, raceme-like, few-flowered.
Flowers Actinomorphic. Pedicels articulated. Hypanthium well developed. Half epigyny (in reality hypogyny with gynoecium sunken into receptacle). Sepals five, with imbricate quincuncial aestivation, whorled, connate, persistent (in Gerrardina foliosa three sepals larger than remaining two sepals). Petals five, with imbricate quincuncial aestivation, whorled, thin, often clawed, somewhat connate, alternating with sepals. Nectariferous disc cupular, quinquelobate, with lobes alternating with stamens.
Androecium Stamens five, obhaplostemonous, antepetalous, alternisepalous. Filament connate into tubular, infundibuliform, cup-shaped or spherical structure, free from tepals. Anthers usually basifixed, non-versatile?, tetrasporangiate, introrse, longicidal (dehiscing by longitudinal slits). Tapetum secretory? Staminodia absent.
Pollen grains Microsporogenesis simultaneous? Pollen grains tricolpor(oid)ate, subangulaperturate, shed as monads, ?-celled at dispersal. Exine semitectate, with columellate infratectum, finely reticulate.
Gynoecium Pistil composed of two connate carpels. Ovary superior, unilocular. Style single, simple. Stigma subcapitate or somewhat bifid, type? Pistillodium absent.
Ovules Placentation apical. Ovules two per carpel, anatropous, pendulous, bitegmic?, crassinucellar? Micropyle ?-stomal. Outer integument ? cell layers thick. Inner integument ? cell layers thick. Megagametophyte monosporous, Polygonum type? Endosperm development? Endosperm haustoria? Embryogenesis?
Fruit A one- to four-seeded, pendant, dry or fleshy berry, with persistent calyx.
Seeds Seed ellipsoid to plano-convex. Aril absent. Raphe protruding. Testa glabrous, finely reticulate. Sarcotesta present (exotesta?). Ribbon-like laterally compressed cells with lignified walls (endotesta?) present inside sarcotesta. Tegmen? Perisperm not developed. Endosperm? Embryo minute, straight, chlorophyll? Cotyledons two. Germination?
Cytology n = ?
DNA
Phytochemistry Virtually unknown. Cyanogenic glycosides not found.
Use Unknown.
Systematics Gerrardina (2; G. eylesiana: tropical East Africa; G. foliosa: southernmost and southeasternmost Africa).
Gerrardina is probably sister to Petenaea (Petenaeaceae).
PETENAEACEAE Christenhusz, M. F. Fay & M. W. Chase 2010 |
Genera/species 1/1
Distribution Central America.
Fossils Unknown.
Habit Bisexual, shrubs or small trees (sometimes lianas). Flowers apetalous, with numerous long pink moniliform hairs.
Vegetative anatomy Phellogen? Vessels present in radial multiples. Vessel elements with simple perforation plates; lateral pits alternate or scalariform, simple pits. Non-vestured pits present. Imperforate tracheary xylem elements fibre tracheids or libriform fibres, septate. Wood rays uniseriate to multiseriate, heterocellular. Axial parenchyma rare or absent. Sieve tube plastids ? type. Nodes 3:3? Crystals?
Trichomes Hairs unicellular or multicellular, uniseriate or branched.
Leaves Alternate (spiral), simple, entire, with ? ptyxis. Stipules minute; leaf sheath absent. Petiole vascular bundle transection? Venation palmate. Stomata? Cuticular wax crystalloids? Leaf margin finely serrate.
Inflorescence Terminal, cymose panicle.
Flowers Actinomorphic. Hypogyny. Sepals four (or five), with valvate aestivation, whorled, free, with basal adaxial glands. Numerous pink moniliform hairs present on adaxial side of sepals. Petals absent. Nectariferous disc annular, with glanduliferous lobes alternating with stamens.
Androecium Stamens eight to twelve, obdiplostemonous, alternisepalous. Filaments connate into a tubular, infundibuliform, cup-shaped or spherical structure, free from tepals. Anthers dorsifixed, non-versatile, tetrasporangiate, latrorse, longicidal (dehiscing by apical slit opening along anther). Tapetum secretory? Staminodia absent.
Pollen grains Microsporogenesis simultaneous? Pollen grains tricolpor(oid)ate, subangulaperturate, shed as monads, ?-celled at dispersal. Exine semitectate, with columellate? infratectum, finely reticulate.
Gynoecium Pistil composed of four or five connate carpels. Ovary superior, unilocular. Style single, simple, long. Stigma subcapitate or somewhat bifid, type? Pistillodium absent.
Ovules Placentation axile. Ovules numerous per carpel, ?-tropous, bitegmic?, crassinucellar? Micropyle ?-stomal. Outer integument ? cell layers thick. Inner integument ? cell layers thick. Megagametophyte monosporous, Polygonum type? Endosperm development? Endosperm haustoria? Embryogenesis?
Fruit A many-seeded lobed berry, with persistent style.
Seeds Aril absent. Testa? Exotestal cells? Endotesta? Tegmen? Perisperm not developed. Endosperm? Embryo?, chlorophyll? Cotyledons two. Germination?
Cytology n = ?
DNA
Phytochemistry Unknown.
Use Unknown.
Systematics Petenaea (1; P. cordata; southern Mexico, Guatemala, Belize, Honduras).
Petenaea appears to be sister to Gerrardina (Gerrardinaceae).
TAPISCIACEAE (Pax) Takht. |
Huerteaceae Doweld in Byull. Mosk. Obshch. Ispyt. Prir., Biol. 105(5): 59. 9 Oct 2000
Genera/species 2/5–6
Distribution Southern and southeastern China, northern Vietnam, the West Indies, northern Andes.
Fossils Fossil fruits and seeds of Tapiscia are known from mid-Eocene layers in Oregon and from the Eocene of western Europe.
Habit Bisexual or unisexual (male; andromonoecious), evergreen (Huertea) or deciduous (Tapiscia) trees.
Vegetative anatomy Phellogen? Vessel elements with scalariform perforation plates; lateral pits scalariform, opposite or alternate, simple and/or bordered pits. Imperforate tracheary xylem elements fibre tracheids (Huertea) or libriform fibres (Tapiscia) with simple and bordered pits, usually septate (sometimes non-septate). Wood rays multiseriate, heterocellular. Axial parenchyma absent. Wood fluorescent. Sieve tube plastids? Nodes 3:3, 4:4 or 5:5, tri-, tetra- or pentalacunar with three, four or five leaf traces, respectively. Prismatic calciumoxalate crystals abundant in Huertea.
Trichomes Hairs unicellular or multicellular, uniseriate.
Leaves Alternate (spiral), imparipinnate or trifoliolate, articulated (with stipels or glands at joint), with ? ptyxis. Stipules usually present (cauline?), caducous (in Huertea sometimes absent); leaf sheath absent. Petiole vascular bundle transection annular. Venation pinnate. Stomata paracytic. Cuticular wax crystalloids? Domatia often present on abaxial side of lamina in axils of lateral veins and along these. Mucilage cells present. Small ‘glands’ present on petiole base, and at points of articulation between petiolules and rhachis. Leaflet margins serrate.
Inflorescence Axillary, spike-like panicle (often with spike-like branches).
Flowers Actinomorphic, small. Hypanthium usually short. Hypogyny. Sepals five (or six), with imbricate quincuncial (Tapiscia) or valvate (Huertea) aestivation, free (Huertea) or connate at base into tube (Tapiscia). Petals five (or six), with imbricate quincuncial (Tapiscia) aestivation, free. Nectariferous disc absent in Tapiscia, in Huertea intrastaminal, quinquelobate with alternipetalous lobes.
Androecium Stamens five (or six), haplostemonous, antesepalous, alternipetalous. Filaments free from each other and from tepals. Anthers dorsifixed (Tapiscia), versatile?, tetrasporangiate, introrse (Tapiscia) or extrorse (Huertea), longicidal (dehiscing by longitudinal slits). Tapetum secretory? Staminodia absent?
Pollen grains Microsporogenesis simultaneous? Pollen grains tricolp(or)ate, shed as monads, ?-cellular at dispersal. Exine tectate or semitectate, with columellate infratectum, reticulate (Huertea) or microreticulate to perforate (Tapiscia).
Gynoecium Pistil composed of two connate carpels. Ovary superior, unilocular, in Huertea with basal septum. Style single, simple (Tapiscia) or bifid (Huertea), with canals. Stigma single, bilobate (Tapiscia) or stigmas two, punctate (Huertea), type? Pistillodium?
Ovules Placentation axile or basal-axile (Huertea) or basal to subbasal (Tapiscia). Ovule one per carpel (Huertea) or one per ovary (Tapiscia), anatropous, erect, apotropous, bitegmic, crassinucellar. Micropyle ?-stomal. Outer integument ? cell layers thick. Inner integument ? cell layers thick. Hypostase present (Tapiscia). Megagametophyte monosporous, Polygonum type? Endosperm development? Endosperm haustoria? Embryogenesis?
Fruit A drupe (Huertea) or a drupaceous berry (Tapiscia).
Seeds Aril absent. Testa thin-walled. Exotesta? Mesotesta in Huertea with sclerotic cells. Exotegmen fairly massive, fibrous. Endotegmen? Chalaza in Tapiscia vascularized with tracheids and curved inwards into endosperm. Perisperm not developed. Endosperm copious, horny, oily. Embryo small to medium-sized, straight, chlorophyll? Cotyledons two, flattened. Germination?
Cytology n = 13, 15
DNA
Phytochemistry Unknown.
Use Timber.
Systematics Tapiscia (1–2; T. sinensis, T. yunnanensis; southern and southeastern China, northern Vietnam), Huertea (4; H. cubensis, H. glandulosa, H. granadina, H. putumayensis; the West Indies, the Andes in Colombia to Peru).
Tapisciaceae are sister-group to Dipentodontaceae (Dipentodon and Perrottetia) or possibly to Perrottetia.
Literature
Alford MH. 2006. Gerrardinaceae: a new family of African flowering plants unresolved among Brassicales, Huerteales, Malvales, and Sapindales. – Taxon 55: 959-964.
Carlquist S, Hoekman DA. 1985. Wood anatomy of Staphyleaceae: ecology, statistical correlations, and systematics. – Flora 177: 195-216.
Chen F-J, Zhao Z-G, Liang H-W, Xiong D, Li F-L. 2007. Chromosome number variation of cells in reiterative subcultures of embryonic callus of Tapiscia sinensis Oliv. – Acta Bot. Boreali-Occid. Sin. 27: 1600-1604.
Christenhusz MJM, Fay MF, Clarkson JJ, Gasson P, Morales Can J, Jiménez Barrios JB, Chase MW. 2010. Petenaeaceae, a new angiosperm family in Huerteales with a distant relationship to Gerrardina (Gerrardinaceae). – Bot. J. Linn. Soc. 164: 16-25.
Cuatrecasas J. 1953. Huertea, un genre nouveau pour la flore de Colombie. – Bull. Soc. Bot. France 100: 159-163.
Dickison WC. 1986. Floral morphology and anatomy of Staphyleaceae. – Bot. Gaz. 147: 312-326.
Dickison WC. 1987a. Leaf and nodal anatomy and systematics of Staphyleaceae. – Bot. Gaz. 148: 475-489.
Dickison WC. 1987b. A palynological study of the Staphyleaceae. – Grana 26: 11-24.
Ding Hou. 1962. Celastraceae I. – In: Steenis CGGJ van (ed), Flora Malesiana I, 6, Wolters-Noordhoff, Groningen, pp. 227-291.
Dunn ST. 1911. Dipentodon: a new genus of uncertain systematic position. – Kew Bull. 1911: 310-313.
Fischer CEC. 1941. Contributions to the Flora of Burma XVIII. – Kew Bull. 1940: 288-294.
Hartog RM den, Baas P. 1978. Epidermal characters of the Celastraceae sensu lato. – Acta Bot. Neerl. 27: 355-388.
Jin Q-J, Wei Z-X. 2002. Studies on pollen morphology of Stachyuraceae and Staphyleaceae. – Acta Bot. Yunnan. 24: 57-63. [In Chinese]
Krause J. 1942. Staphyleaceae. – In: Engler A (†), Harms H, Mattfeld J (eds), Die natürlichen Pflanzenfamilien, 2. Aufl., Bd. 20b, W. Engelmann, Leipzig, pp. 255-321.
Kubitzki K. 2002. Tapisciaceae. – In: Kubitzki K, Bayer C (eds), The families and genera of vascular plants V. Flowering plants. Dicotyledons. Malvales, Capparales and non-betalain Caryophyllales, Springer, Berlin, Heidelberg, New York, pp. 369-370.
Kukachka BF. 1962. Wood anatomy of Petenaea cordata Lundell (Elaeocarpaceae). – Wrightia 3: 36-40.
Liu J-S, Cheng J-R. 1989. Comparative anatomy of wood in the genus Dipentodon. – J. Sichuan For.Sci. Techn. 10: 9-13. [In Chinese]
Liu J-S, Cheng J-R. 1991. On the systematic position of genus Dipentodon Dunn. – J. Wuhan Bot. Res. 9: 29-39. [In Chinese]
Liu W-Z, Kang H-Q, Zheng H-C, Feng Y-Z. 2008. An investigation on the sexual reproductive cycle in Tapiscia sinensis. – J. Syst. Evol. 46: 175-182.
Lobreau-Callen D. 1982. Structures et affinités polliniques des Cardiopterygaceae, Dipentodontaceae, Erythropalaceae et Octoknemataceae. – Bot. Jahrb. Syst. 103: 371-412.
Loesener T. 1942. Celastraceae. – In: Engler A (†), Harms H, Mattfeld J (eds), Die natürlichen Pflanzenfamilien, 2. Aufl., Bd. 20b, W. Engelmann, Leipzig, pp. 87-197.
Lundell CL. 1962. Plantae Mayanae V. Petenaea cordata, a new genus and species in the Elaeocarpaceae, and other taxonomic notes. – Wrightia 3: 21-35.
Ma J-S, Bartholomew B. 2008. Dipentodontaceae. – In: Wu Z-Y, Raven PH (eds), Flora of China, Science Press, Beijing & Missouri Botanical Garden Press, St. Louis, vol. 11, pp. 494-495.
Manchester SR. 1988. Fruits and seeds of Tapiscia (Staphyleaceae) from the Middle Eocene of Oregon, USA. – Tertiary Res. 9: 59-66.
Matthews ML, Endress PK. 2005. Comparative floral structure and systematics in Celastrales (Celastraceae, Parnassiaceae, Lepidobotryaceae). – Bot. J. Linn. Soc. 149: 129-194.
Merrill ED. 1941. The Upper Burma plants collected by Captain F. Kingdon Ward on the Vernay-Cutting expedition, 1938-39. – Brittonia 4: 20-188.
Milne-Redhead E. 1939. Gerrardina eylesiana Milne-Redhead. – Hooker’s Icon. Pl., 5th Ser. 4: 1-2, t. 3390.
Oliver D. 1870. Gerrardina foliosa, Oliv. – Hooker’s Icon. Pl., 3rd Ser., 1: 60, t. 1075.
Oliver D. 1890. Tapiscia sinensis. – Hooker’s Icon. Pl., 3rd Ser., 5, t. 1928.
Pax F. 1896. Staphyleaceae. – In: Engler A, Prantl K (eds), Die natürlichen Pflanzenfamilien III(5), W. Engelmann, Leipzig, pp. 258-262.
Peng H. 2001. Lectotypification of the name Dipentodon sinicus (Dipentodontaceae). – Taxon 50: 1177-1178.
Peng Y, Chen Z, Gong X, Zhong Y, Shi S. 2003. Phylogenetic position of Dipentodon sinicus: evidence from DNA sequences of chloroplast rbcL, nuclear ribosomal 18S, and mitochondrial matR genes. – Bot. Bull. Acad. Sin. 44: 217-222.
Solereder H. 1892. Über die Staphyleaceengattung Tapiscia Oliv. – Ber. Deutsch. Bot. Ges. 10: 545-551.
Stevens PF. 2001 onwards. Angiosperm Phylogeny Website. Version 10, April 2012 [and more or less continuously updated since] http://www.mobot.org/MOBOT/research /APweb/
Teng L, Liu W-Z. 2009. Passing winter strategies of Tapiscia sinensis during its extremely long reproductive cycle. – J. Wuhan Bot. Res. 27: 70-75. [In Chinese]
Wei Z-X, Jin Q-J, Hong W, Tian X, Chen S-K. 2002. Pollen morphology of Stachyuraceae and related taxa. – Acta Bot. Yunnan. 24: 483-496. [In Chinese]
Worberg A, Alford MH, Quandt D, Borsch T. 2009. Huerteales sister to Brassicales plus Malvales, and newly circumscribed to include Dipentodon, Gerrardina, Huertea, Perrottetia and Tapiscia. – Taxon 58: 468-478.
Yang J, Wu J-C, Gu Z-J. 2009. Karyomorphology of three species in Dipentodon (Dipentodontaceae), Perrottetia (Celastraceae), and Tapiscia (Tapisciaceae) of the order Huerteales and their phylogenetic implications. – J. Syst. Evol. 47: 291-296.
Yuan Q-J, Zhang Z-Y, Peng H, Ge S. 2008. Chloroplast phylogeny of Dipentodon (Dipentodontaceae) in southwestern China and northern Vietnam. – Mol. Ecol. 17: 1054-1065.
Zhang J, Li Z, Fritsch PW, Tian H, Yang A, Yao X. 2015. Phylogeography and genetic structure of a Tertiary relict tree species, Tapiscia sinensis (Tapisciaceae): implications for conservation. – Ann. Bot. 116: 727-737.
Zhang L-B, Simmons MP. 2006. Phylogeny and delimitation of the Celastrales inferred from nuclear and plastid genes. – Syst. Bot. 31: 122-137.
Zhang X-Y, Gao X-Z. 1995. Anatomical studies on secondary xylem and secondary phloem of Dipentodon sinicus. – Acta Bot. Sin. 37: 534-538. [In Chinese with English summary]
Zhou X-J, Ma L, Liu W-Z. 2016. Functional androdioecy in the rare endemic tree Tapiscia sinensis. – Bot. J. Linn. Soc. 180: 504-514.