BRUNIALES Bercht. et J.
Presl
Presl, Přir. Rostlin: 226. Jan-Apr 1820
[‘Bruniaceae’]
BRUNIACEAE R. Br. ex DC.
de Candolle, Prodr. 2: 43. Nov 1825, nom.
cons.
Berzeliaceae Nakai,
Chosakuronbun Mokuroku [Ord. Fam. Trib. Nov.]: 241. 20 Jul 1943;
Brunianae Doweld, Tent. Syst. Plant. Vasc.: liii. 23
Dec 2001
Genera/species 6/81
Distribution Southern coastal
and mountain areas in South Africa (almost restricted to Western and Eastern
Cape), one species of Brunia in southern KwaZulu-Natal.
Fossils The fossilized flowers
of Actinocalyx from the Santonian to the Campanian of Sweden are
similar to those in Bruniaceae.
Habit Bisexual, evergreen
ericoid shrubs (rarely trees). Lignotubers present or absent.
Vegetative anatomy Phellogen
ab initio superficial. Vessel elements with scalariform perforation plates;
lateral pits scalariform to opposite, simple or bordered pits. Imperforate
tracheary xylem element tracheids (in some species of Berzelia and
Staavia with crassulae, thickenings of primary walls between tracheids
and vessels or among tracheids) with bordered pits, non-septate? Wood rays
uniseriate or multiseriate, heterocellular. Axial parenchyma apotracheal
diffuse (rarely diffuse-in-aggregates; sometimes paratracheal scanty
vasicentric). Sieve tube plastids S type. Nodes 1:1, unilacunar with one leaf
trace. Amorphous dark-staining material present in wood of all species.
Rhomboidal calciumoxalate crystals usually present (in wood rays, sometimes in
axial parenchyma); druses present.
Trichomes Hairs unicellular or
multicellular, simple.
Leaves Alternate (spiral,
usually pentastichous), simple, entire, usually ericoid, with ? ptyxis.
Stipules minute or absent (in Staavia resembling colleters); leaf
sheath absent. Venation parallel, usually with three (rarely five to 20) veins,
or leaves one-veined. Stomata usually anomocytic (sometimes cyclocytic, with
transverse orientation), sometimes on adaxial side of lamina only. Cuticular
wax crystalloids as platelets, scales or rods. Mesophyll cells tanniniferous.
Rhomboidal crystals frequent; druses present in mesophyll cells in some
species. Leaf margin entire, often inrolled. Leaf apex with localized phellogen
(cork cells formed from local phellogen), often black, glanduliferous.
Inflorescence Terminal or
axillary, usually headlike (sometimes spike-like), these sometimes in panicles
or raceme-like partial inflorescences, sometimes (Staavia) in
pseudanthia with involucrum consisting of several bracts (in Brunia
subgenus Pseudobaeckia solitary along branches; in Thamnea
solitary terminal).
Flowers Usually actinomorphic
(sometimes slightly zygomorphic), usually small. Usually epigyny or half
epigyny (in Brunia subgenus Raspalia hypogyny). Sepals (four
or) five, with imbricate aestivation, persistent, free or connate at base.
Petals (four or) five, with imbricate aestivation, often clawed, persistent or
caducous, free or more or less connate (initially free), in lower part with
non-vascularized adaxial ridges or swellings. Nectaries intrastaminal,
gynoecial (on upper part of ovary), often as nectariferous disc, or absent.
Androecium Stamens (four or)
five, haplostemonous, antesepalous, alternipetalous, often persistent; abaxial
stamens larger than the remainder. Filaments free from each other, usually free
from petals (sometimes adnate to petals in lower part [epipetalous]). Anthers
dorsifixed, inflexed in bud, usually versatile, tetrasporangiate, introrse,
longicidal (dehiscing by longitudinal slits); connective sometimes prolonged at
apex. Tapetum secretory? Staminodia absent.
Pollen grains
Microsporogenesis simultaneous? Pollen grains tricolpor(oid)ate or
6–10(–11)-colpor(oid)ate, shed as monads, ?-cellular at dispersal. Exine
tectate or semitectate, with columellate infratectum, imperforate, perforate or
reticulate, foveolate, psilate, granulate, regulate, or verrucate.
Gynoecium Pistil composed of
usually two (rarely three to five) connate carpels (in Berzelia and
Brunia subgenus Mniothamnea at least seemingly one carpel).
Ovary usually inferior or semi-inferior, usually bilocular (rarely monomerous
or pseudomonomerous? or trilocular to quinquelocular). Stylodia usually two
(rarely three to five), almost free to nearly entirely connate (style rarely
single, simple). Stigmas capitate, papillate?, Wet type. Pistillodium
absent.
Ovules Placentation
apical-axile. Ovules (one or) two to four (to twelve) per carpel (when a single
carpel, then one ovule), usually anatropous (sometimes pleurotropous),
pendulous, epitropous, unitegmic, tenuinucellar to weakly crassinucellar.
Micropyle long. Integument massive, ? cell layers thick. Endothelium present.
Hypostase present. Megagametophyte monosporous, Polygonum type.
Endosperm development? Endosperm haustoria? Embryogenesis?
Fruit A schizocarp usually
with two one-seeded nutlike or follicle-like mericarps, a nutlet or a
two-seeded loculicidal capsule (sometimes also adaxially dehiscent) with
persistent and sometimes accrescent calyx.
Seed: Seed sometimes with
aril. Exotesta thin. Endotesta? Perisperm not developed. Endosperm copious,
fleshy. Embryo small, straight, well differentiated, chlorophyll? Cotyledons
two. Germination phanerocotylar.
Cytology n = 10, 11, 20–23
– Polyploidy occurring in Berzelia.
DNA
Phytochemistry Insufficiently
known. Flavonols (quercetin, myricetin, isorhamnetin) and proanthocyanidins
(prodelphinidins) present. Ellagic acid not found. Iridoids?
Use Ornamental plants.
Systematics The sister-group
relationships of Bruniaceae
are not entirely clarified, yet they may be sister to Columelliaceae.
Linconia is sister to the remaining Bruniaceae, and the
[Audouinia+Thamnea] clade is sister to the clade [Staavia+[Berzelia+Brunia]]
(Quint & Claßen-Bockhoff 2006a; Claßen-Bockhoff & al. 2011).
Linconieae Quint et
Class.-Bockh. in Taxon 60: 1144. 4 Aug 2011
1/3. Linconia (3; L.
alopecuroidea, L. cuspidata, L. ericoides; Western Cape,
southwestern Eastern Cape). Lignotubers present. Each stoma surrounded by
cuticular rim. Petals stiff and inflexible. Anthers sagittate, with thecae
apically connate and with apex ending in sterile conspicuous tip. n = ?
[Audouinieae+Brunieae]
Petals flexible. Anthers without sterile
apex.
Audouinieae Nied. in
H. G. A. Engler et K. A. E. Prantl, Nat. Pflanzenfam. III, 2a: 133. 9 Mar
1891
2/14. Audouinia (5; A.
capitata, A. esterhuyseniae, A. hispida, A.
laevis, A. laxa; Western Cape), Thamnea (9; Western
Cape). – Lignotubers present. Stomata often surrounded by cuticular rim.
Anthers without sterile apex, with thecae connate along their entire
length.Audouinia with thecae entirely adnate to connective and with
ovary usually trilocular. n = 11 (Audouinia).
Brunieae Quint et
Class.-Bockh. in Taxon 60: 1146. 4 Aug 2011
3/64. Staavia
(11; Western Cape), Berzelia
(16; Western and Eastern Cape), Brunia
(37; Western Cape, southern Eastern Cape, one species in KwaZulu-Natal). –
Western and Eastern Cape, KwaZulu-Natal. Lignotubers present or absent. Stomata
not surrounded by cuticular rim. Anthers versatile, without sterile apex.
Petals and filaments in Brunia
subgenus Lonchostoma secondarily postgenitally connate. – Staavia
is sister to [Berzelia+Brunia].
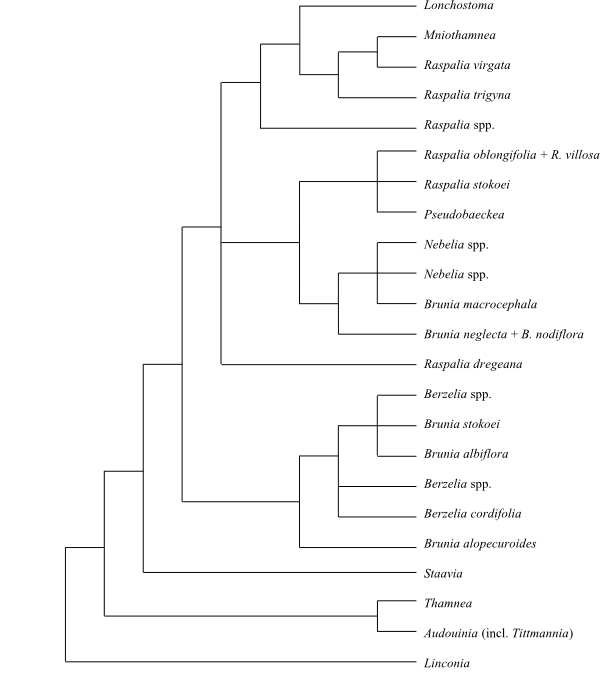 |
Cladogram of Bruniaceae based on DNA
sequence data (Quint & Claßen-Bockhoff 2006a).
|
Don in Edinburgh New Philos. J. 6: 46, 49.
Oct-Dec 1828 [’Columellieae’], nom. cons.
Columelliales D. Don
in C. F. P. von Martius, Consp. Regn. Veg.: 27. Sep-Oct 1835
[’Columellieae’]; Desfontainiaceae
(Endl.) L. K. G. Pfeiffer, Nomencl. Bot. 1: 1037. 21 Feb 1873
[’Desfontaineae’], nom. cons.
Desfontainiales Takht., Divers. Classif. Fl. Pl.:
377. 24 Apr 1997
Genera/species 2/8
Distribution Southern Mexico
to Cape Horn in Chile.
Fossils Unknown.
Habit Bisexual, evergreen
shrubs or small trees. Bud scales absent.
Vegetative anatomy Phellogen
ab initio pericyclic (in Desfontainia near phloem). Primary medullary
rays in Desfontainia very thin, mostly uniseriate. Medulla
parenchymatous (Desfontainia). Pericyclic fibres absent. Vessel
elements with scalariform perforation plates; lateral pits scalariform
(Desfontainia), or alternate or absent (Columellia), simple
or bordered pits. Vestured pits absent. Imperforate tracheary xylem elements
tracheids or fibre tracheids with simple or bordered pits, non-septate (also
vasicentric or vascular tracheids). Wood rays uniseriate, heterocellular. Axial
parenchyma apotracheal usually diffuse, or paratracheal vasicentric
(Columellia). Intraxylary phloem absent. Sieve tube plastids S type.
Nodes 1:1, unilacunar with one leaf trace (Columellia). Pericycle with
scattered sclereids (Desfontainia). Calciumoxalate as druses.
Trichomes Hairs simple
(sometimes absent in Desfontainia); glands and glandular hairs present
or absent.
Leaves Opposite, simple,
entire, in Columellia strongly asymmetrical, in Desfontainia
coriaceous, with conduplicate ptyxis. Stipules small interpetiolar to
intrapetiolar (Desfontainia) or absent (Columellia, sometimes
in Desfontainia); leaf sheath absent. Colleters present in
Desfontainia. Petiole vascular bundle transection arcuate; central
vascular bundle largest (Columellia). Abaxial side of lamina in
Columellia sometimes glanduliferous. Venation pinnate. Stomata
anomocytic. Cuticular wax crystalloids in at least Columellia as
tubuli (scarcely branched), dominated by β-diketones. Mesophyll in
Columellia with calciumoxalate druses. Leaf margin entire to serrate,
usually with glandular tip (Columellia) or serrate-dentate to
spinose-serrate (Desfontainia).
Inflorescence Terminal?
(Columellia) or axillary? (Desfontainia), few-flowered cymes
or flowers solitary (in Desfontainia usually solitary).
Flowers Actinomorphic
(Desfontainia) or somewhat zygomorphic (Columellia,
Desfontainia), medium-sized. Epigyny (Columellia) or hypogyny
(Desfontainia). Sepals (four or) five (to eight), with valvate or
somewhat imbricate aestivation, persistent, usually more or less connate in
lower part; median sepal abaxial. Petals (four or) five (to eight), with
imbricate (Columellia) or imbricate to contorted
(Desfontainia) aestivation, connate at base (Columellia) or
connate (petals possibly initially free), tubular and fleshy
(Desfontainia). Nectary absent. Disc absent.
Androecium Stamens in
Columellia two (adaxial pair), alternating with adaxial and lateral
calyx lobes, alternipetalous; in Desfontainia five, antesepalous,
alternipetalous. Filaments short, stout, free, in Columellia inserted
at petal base, in Desfontainia adnate to uppermost part of corolla
tube. Anthers dorsifixed (Columellia) or basifixed
(Desfontainia), non-versatile, tetrasporangiate, latrorse to extrorse
(Columellia) or introrse (Desfontainia), longicidal
(dehiscing by longitudinal slits); thecae semicircular and attached their
entire length to expanded connective (Columellia), or embedded in
connective (Desfontainia); connective well developed, in
Columellia expanded at apex. Tapetum amoeboid-periplasmodial
(Columellia) or secretory (Desfontainia). Staminodia
absent.
Pollen grains
Microsporogenesis simultaneous. Pollen grains tricolporate, shed as monads,
tricellular at dispersal (Columellia). Exine tectate or semitectate,
with columellate infratectum, finely reticulate (Columellia) or
perforate (Desfontainia), rugulate.
Gynoecium Pistil composed of
two median (Columellia) or (three to) five (to seven) alternisepalous,
antepetalous (Desfontainia) connate carpels. Ovary inferior
(Columellia) or superior (Desfontainia), incompletely
bilocular (Columellia), or bilocular to quinquelocular at base and
unilocular at apex (Desfontainia). Style single, simple, short and
thick (Columellia), or longer and filiform, persistent
(Desfontainia). Stigma almost cup-shaped, bilobate or quadrilobate
(Columellia), or capitate to somewhat quinquelobate
(Desfontainia), type? Pistillodium absent.
Ovules Placentation
intrusively parietal to axile; placentae in Columellia forming
secondary incomplete septa; placentation in Desfontainia mainly axile
yet strongly intrusively parietal in uppermost part. Ovules numerous per
carpel, anatropous, ascending (Columellia), unitegmic, tenuinucellar
(Columellia) or crassinucellar (Desfontainia). Integument six
to nine cell layers thick (Desfontainia). Parietal tissue possibly
absent (Columellia). Haustorial suspensor large in
Desfontainia. Endothelium poorly developed (Desfontainia).
Megagametophyte monosporous, Polygonum type. Endosperm development in
Columellia ?, in Desfontainia cellular. Endosperm haustoria
in Columellia ?, in Desfontainia chalazal and micropylar.
Embryogenesis solanad (Desfontainia).
Fruit A septicidal and
partially loculicidal capsule (Columellia) or a berry
(Desfontainia) with numerous seeds and persistent calyx.
Seeds Aril? Seed coat
exotestal. Exotestal cells elongate (Columellia) or polygonal
(Desfontainia); anticlinal exotestal cells strongly thickened, with
plasmodesmata; outer walls of exotestal cells pectic and remaining cell walls
lignified (Desfontainia). Endotesta? Perisperm not developed.
Endosperm copious (in Desfontainia starchy). Embryo small, straight,
well differentiated, without chlorophyll. Cotyledons two. Germination?
Cytology n = 7
(Desfontainia)
DNA
Phytochemistry Insufficiently
known. Route I iridoids (secoiridoids: loganin, loganic acid, etc.) and
cornoside present in Desfontainia. Proanthocyanidins, saponins and
cyanogenic compounds not found. Tannins not found in Desfontainia.
Myricetin?
Use Medicinal plants,
carpentry, dyeing substances.
Systematics
Columellia (5; C. lucida, C. oblonga, C.
obovata, C. subsessilis, C. weberbaueri; the Andes from
southern Colombia to Bolivia), Desfontainia (3; D. fulgens:
Chile, western Argentina; D. spinosa: the Andes from Costa Rica to
Cape Horn in Chile; D. splendens: mountains from southern Mexico to
Bolivia).
The sister-group relationship of Columelliaceae is not entirely
clarified, although they seem to be sister to Bruniaceae.
Literature
Bremer B, Olmstead RG, Struwe L, Sweere JA.
1994. rbcL sequences support exclusion of Retzia,
Desfontainia, and Nicodemia from the Gentianales.
– Plant Syst. Evol. 190: 213-230.
Brizicky GK. 1961. A synopsis of the genus
Columellia (Columelliaceae). – J. Arnold
Arbor. 42: 363-372.
Carlquist SJ. 1978. Wood anatomy of Bruniaceae: correlations with ecology,
phylogeny, and organography. – Aliso 9: 323-364.
Carlquist SJ. 1990. Leaf anatomy of Geissolomataceae
and Myrothamnaceae as a
possible indicator of relationship to Bruniaceae. – Bull. Torrey Bot. Club
117: 420-428.
Carlquist SJ. 1991. Leaf anatomy of Bruniaceae: ecological, systematic, and
phylogenetic aspects. – Bot. J. Linn. Soc. 107: 1-34.
Claßen-Bockhoff R. 2000. Inflorescences in
Bruniaceae, with general comments on
inflorescences in woody plants. – Opera Bot. Belg. 12: 5-310.
Claßen-Bockhoff R, Oliver EGH, Hall AV,
Quint M. 2011. A new classification of the South African endemic family Bruniaceae based on molecular and
morphological data. – Taxon 60: 1138-1155.
Dahlgren RMT, Wyk AE van. 1988. Structures
and relationships of families endemic to or centered in southern Africa. –
Monogr. Syst. Bot. Missouri Bot. Gard. 25: 1-94.
Daniel M, Sabnis SD. 1979. Chemotaxonomy of
Loganiaceae. – Curr. Sci.
48: 383-385.
Dümmer RA. 1912. An enumeration of the Bruniaceae. – J. Bot. 50 [Suppl. 2]:
1-37.
Fagerström K. 1975. 182. Columelliaceae. – In: Harling G,
Sparre B (eds), Flora of Ecuador 4, Swedish Natural Science Research Council,
Stockholm, pp. 3-5.
Fritsch K. 1895. Columelliaceae. – In: Engler A,
Prantl K (eds), Die natürlichen Pflanzenfamilien IV(3b), W. Engelmann,
Leipzig, pp. 186-188.
Goldblatt P. 1981. Chromosome cytology of Bruniaceae. – Ann. Missouri Bot. Gard.
68: 546-550.
Gregory M. 1998. Columelliaceae. – In: Cutler DF,
Gregory M (eds), Anatomy of the dicotyledons, 2nd ed., IV, Clarendon
Press, Oxford, pp. 117-120.
Gustafsson MHG, Bremer K. 1995. Morphology
and phylogenetic interrelationships of the Asteraceae, Calyceraceae, Campanulaceae, Goodeniaceae, and
related familes (Asterales). – Amer. J. Bot. 82: 250-265.
Hall AV. 1987. Evidence of a Cretaceous
alliance for the Bruniaceae. –
South Afr. J. Sci. 83: 58-59.
Hall AV. 1988. Systematic palynology of the
Bruniaceae. – Bot. J. Linn. Soc.
96: 285-296.
Hall AV, Oliver EGH, Claßen-Bockhoff R.
2010. Bruniaceae: new species of
Thamnea and Brunia from western Cape, South Africa. –
Bothalia 40: 96-101.
Hasselberg GBE. 1937. Zur Morphologie des
vegetativen Sproßes der Loganiaceen. – Symb. Bot. Upsal. 2(3): 1-170.
Hoc P, Bravo L. 1984. Estudio palinológico
sobre las especies presentes en Argentina de Spigelia,
Strychnos, y Desfontainia (Loganiaceae). – Kurtziana
17: 71-89.
Houghton PJ, Ming L-L. 1985. Iridoids from
Desfontainia spinosa. – Phytochemistry 24: 1841-1842.
Houghton PJ, Ming L-L. 1986a. Iridoids,
iridoid-triterpenoid congeners and lignans from Desfontainia spinosa.
– Phytochemistry 25: 1907-1912.
Houghton PJ, Ming L-L. 1986b. Triterpenoids
from Desfontainia spinosa. – Phytochemistry 25: 1939-1944.
Jay MM. 1968. Distribution des flavonoïdes
chez les Bruniacées. – Taxon 17: 484-488.
Kadereit JW, Bittrich V (eds). 2016. The
families and genera of vascular plants XIV. Flowering plants – eudicots –
Aquifoliales, Boraginales, Bruniales, Dipsacales, Escalloniales, Garryales, Paracryphiales,
Solanales (except Convolvulaceae), Icacinaceae, Metteniusaceae,
Vahliaceae. – Springer,
412 pp.
Kirchner R. 1904. Beiträge zur Kenntniss der
Bruniaceen. – Ph.D. diss., Universität Breslau, Poland.
Lange JHD, Boucher C, Walt JJA van der. 1993.
Autecological studies on Audouinia capitata (Bruniaceae) 3. Pollination biology. –
South Afr. J. Bot. 59: 135-144.
Lange JHD, Walt JJA van der, Boucher C.
1993a. Autecological studies on Audouinia capitata (Bruniaceae) 5. Seed development,
abortion and pre-emergent reproductive success. – South Afr. J. Bot. 59:
156-167.
Lange JHD, Walt JJA van der, Boucher C.
1993b. Autecological studies on Audouinia capitata (Bruniaceae) 6. Nutritional aspects of
the developing ovule. – South Afr. J. Bot. 59: 168-177.
Leeuwenberg AJM. 1969. Notes on American Loganiaceae IV. Revision of
Desfontainia Ruiz et Pav. – Acta Bot. Neerl. 18: 669-679.
Leeuwenberg AJM (ed). 1980. Angiospermae:
Ordnung Gentianales Fam. Loganiaceae. – In: Hiepko P,
Melchior H (eds), Die natürlichen Pflanzenfamilien, 2. Aufl., Vol. 28bI,
Duncker & Humblot, Berlin, pp. 1-255.
Leinfellner W. 1964a. Über die falsche
Sympetalie bei Lonchostoma und anderen Gattungen der Bruniaceen. –
Österr. Bot. Zeitschr. 111: 345-353.
Leinfellner W. 1964b. Sind die Kronblätter
der Bruniaceae peltat gebaut? –
Österr. Bot. Zeitschr. 111: 500-526.
Maldonado de Magnano S. 1986. Estudios
embriológicos en Desfontainia spinosa (Desfontainiaceae). –
Darwiniana 27: 207-224.
Niedenzu F. 1891. Bruniaceae. – In: Engler A, Prantl K
(eds), Die natürlichen Pflanzenfamilien III(2a), W. Engelmann, Leipzig, pp.
131-136.
Niedenzu F, Harms H. 1930. Bruniaceae. – In: Engler A, Harms H
(eds), Die natürlichen Pflanzenfamilien, 2. Aufl., Bd. 18a, W. Engelmann,
Leipzig, pp. 288-303.
Oliver EGH, Oliver IM. 1999. Bruniaceae. A new species of
Linconia from Western Cape. – Bothalia 29: 256-258.
Pillans NS. 1947. A revision of Bruniaceae. – J. South Afr. Bot. 13:
121-206.
Powrie E. 1969a. Types of Bruniaceae in the Thunberg Herbarium.
– J. South Afr. Bot. 35: 327-339.
Powrie E. 1969b. A new species of
Tittmannia (Bruniaceae). –
J. South Afr. Bot. 35: 363-366.
Quint M. 2004. Evolution of Bruniaceae: evidence from molecular and
morphological studies. – Ph.D. diss., Universität Mainz, Germany.
Quint M, Claßen-Bockhoff R. 2006a. Phylogeny
of Bruniaceae based on matK
and ITS sequence data. – Intern. J. Plant Sci. 167: 135-146.
Quint M, Claßen-Bockhoff R. 2006b. Floral
ontogeny, petal diversity and nectary uniformity in Bruniaceae. – Bot. J. Linn. Soc. 150:
459-477.
Quint M, Claßen-Bockhoff R. 2008. Ancient or
recent? Insights into the temporal evolution of the Bruniaceae. – Organisms Divers. Evol.
8: 293-304.
Saxton WT. 1910. The ovule of the Bruniaceae. – Trans. Roy. Soc. South
Africa 2: 27-31.
Schlechter R. 1920. Die Columelliaceae. – Notizbl. Bot.
Gart. Berlin-Dahlem 7: 352-358.
Schultes RE. 1977. De plantes toxicariis e
mundo novo tropicale commentationes: 15. Desfontainia: a new Andean
hallucinogen. – Bot. Mus. Leafl. 25: 99-104.
Schultes RE. 1989. De speciebus
varietatibusque Desfontainia Columbianae notae. – Rev. Acad. Colomb.
Ci. Exact. Fisic. Natur. 17: 313-319.
Scott G. 1999. A chemosystematic and
cladistic study of the Southern African endemic family Bruniaceae. – Ph.D. diss., Department
of Botany, University of Cape Town, Republic of South Africa.
Solereder H. 1895. Loganiaceae. – In: Engler A,
Prantl K (eds), Die natürlichen Pflanzenfamilien IV(2), W. Engelmann, Leipzig,
pp. 19-50.
Stern WL, Brizicky GK, Eyde RH. 1969.
Comparative anatomy and relationships of Columelliaceae. – J. Arnold Arbor.
50: 36-75.
Strid A. 1968. A new species of
Lonchostoma (Bruniaceae).
– Bot. Not. 121: 312-316.
Tieghem P van. 1903. Sur les Columelliacées.
– Ann. Sci. Nat. 17-18: 155-164.
Villiers SE de, Cadman A. 1997. The
palynology of tertiary sediments from a palaeochannel in Namaqualand, South
Africa. – Palaeontol. Afr. 34: 69-99.
Weberling F. 1976. Weitere Untersuchungen zur
Morphologie des Unterblattes bei den Dikotylen IX. Saxifragaceae
s.l., Brunelliaceae,
und Bruniaceae. – Beitr. Biol.
Pflanzen 52: 163-181.
Weigend M. 2001. Desfontainia Ruiz
& Pav. (Desfontainiaceae) revisited – a first step back towards
α-diversity. – Bot. Jahrb. Syst. 123: 281-301.